La levure de boulangerie, Saccharomyces cerevisiae, est un organisme unicellulaire
haplodiplobiontique (cycle haploïde et cycle diploïde de même importance). Sa
multiplication végétative est assurée par le bourgeonnement d’une cellule fille à
partir d’une cellule mère, les deux cellules étant porteuses d’un noyau contenant le
même lot de chromosomes à l’issue de la division nucléaire (mitose).
Deux cellules haploïdes, de signe sexuel différent MATa et MATα, sont capables
de fusionner pour donner une cellule diploïde dont la multiplication végétative suit
les mêmes règles que celles des cellules haploïdes.
L’étalement de cellules sur une boîte de milieu nutritif (au moins une source de
carbone, une source d’azote et des éléments minéraux, plus d’éventuels apports en
cas d’auxotrophie) aboutit à la formation de colonies, clones de cellules issues d’une
cellule initiale.
Ces colonies sont bien individualisées si le nombre de cellules
étalées est faible (quelques dizaines ou centaines); elles sont jointives et forment un
tapis cellulaire continu dès que leur nombre est élevé (plusieurs milliers).
Pour croiser deux souches haploïdes de levure, il faut les mettre en contact sur une
boîte de milieu, de manière à ce que des colonies diploïdes puissent s’y développer
tout en prenant garde qu’aucun des deux parents haploïdes ne puissent s’y multiplier.
Dans ce but, on choisit toujours de croiser entre elles, sur un milieu minimum,
des souches porteuses de mutations d’auxotrophie différentes, de sorte que les
parents ne peuvent s’y développer alors que les diploïdes le peuvent, par complémentation
fonctionnelle.
De ce fait, dans le croisement d’une souche mutante par
une souche SSR, cette dernière est « sauvage » pour le (ou les) gène(s) impliqué(s)
dans le phénotype étudié, mais les deux souches sont obligatoirement porteuses de
mutations d’auxotrophie servant de « marqueurs de sélection des diploïdes ».
Pour étudier les produits de la méiose, il est nécessaire d’induire la sporulation de
cellules diploïdes afin de recueillir les spores haploïdes.
L’analyse génétique des
spores issues de la méiose est directe puisqu’elles sont cultivables.
Ce n’est pas le
cas des gamètes chez les organismes diplobiontiques (cycle haploïde réduit aux
gamètes, drosophile par exemple) où leur analyse génétique est indirectement
réalisée par l’analyse d’une descendance issue du croisement entre le donneur des
gamètes à analyser et un autre parent (F1 × F1 ou test-cross).
Les spores sont obtenues par étalement de cellules diploïdes sur un milieu de
sporulation (pauvre en éléments azotés).
Les cellules diploïdes y arrêtent leur multiplication,
passent en méiose et donnent un asque contenant quatre spores haploïdes
en phase stationnaire.
La dissection de l’asque et l’étalement des spores de chaque asque, sur un milieu
adéquat, permettent à chacune d’initier un développement clonal haploïde sous la
forme de colonies individualisées.
Ce milieu de développement des spores doit contenir tous les éléments autorisant ce
développement, notamment les apports correspondant aux auxotrophies parentales des
marqueurs de sélection des diploïdes, qu’on retrouvera chez certaines des spores, plus
les éléments correspondant éventuellement au(x) gène(s) étudié(s) dans le croisement.
Pour déterminer le génotype des spores haploïdes, il faut tester leur phénotype
d’auxotrophie en repiquant quelques cellules de chaque colonie sur autant de
milieux adéquats qu’il y a d’auxotrophies en jeu. On peut tester le signe sexuel en
testant leur capacité à former des diploïdes avec une souche MATa ou MATα.
Le génotypage du (ou des) gène(s) étudié(s) dans le croisement est assuré par un test
phénotypique spécifique.
Problèmes
:
Problème 10.1
On connaît chez la levure Saccharomyces cerevisiae trois gènes, nommés A,
B et C, dont les mutations peuvent conférer respectivement le phénotype
[ade–] ou [leu–] ou [his–].
Ces trois phénotypes d’auxotrophie sont récessifs.
Le gène A est situé sur le chromosome I, près du centromère, le gène B est
situé sur le chromosome III, près du centromère, le gène C est distant du
gène B de 20 unités de recombinaison.
On suppose qu’il n’y a, au plus
qu’un crossing-over possible entre les locus des gènes B et C.
On croise une souche mutée dans les gènes A et C, de phénotype
[ade–; his–], par une souche mutée dans le gène B, de phénotype [leu–].
1. Quelle est la composition du milieu de culture des deux souches parentales
et celle du milieu de croisement ?
Justifiez votre réponse.
2. Vous ferez le schéma clair de chacun des scénarios possibles de la
méiose, en précisant à chaque fois le génotype et le phénotype des spores
qui en sont issus.
3. Quelles sont les fréquences respectives de chacun de ces scénarios et des
différents types de spores qui en sont issus ?
Solution
1. La souche [ade–; his–] doit être cultivée sur un milieu minimum (Mo) additionné
d’adénine et d’histidine puisqu’elle est auxotrophe pour ces deux molécules; la souche [leu–]
est cultivée sur Mo + leucine.
Mais le croisement peut et doit se faire sur milieu Mo, ainsi les spores haploïdes ne peuvent
développer des colonies et on est sûr que les colonies récupérées sont diploïdes, ayant pu
pousser grâce à la complémentation fonctionnelle pour les trois mutations d’auxotrophie.
Ces mutations servent de « marqueurs de sélection » des diploïdes.
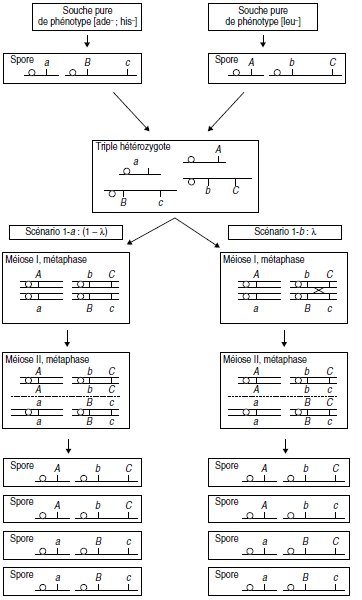
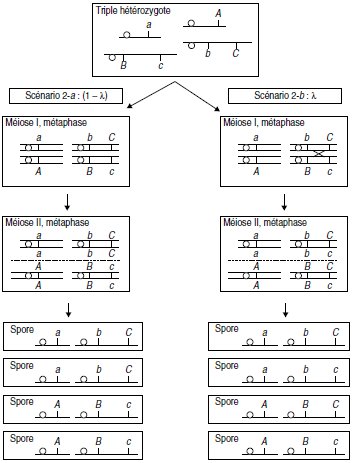
2. La méiose peut suivre quatre scénarios qui correspondent aux diverses dispositions métaphasiques
possibles des gènes entre eux.
Si on considère d’abord les gènes A et B (pris indépendamment de C), on a deux scénarios
possibles de la méiose (fig. 10.1 et fig. 10.2), selon les deux dispositions métaphasiques équifréquentes
correspondant à l’assortiment aléatoire des paires de chromatides non homologues.
Si on considère le gène C par rapport au gène B, chacun des deux scénarios précédents se
subdivise en deux sous-scénarios selon qu’il y a (fig. 10.1 et 10.2, à droite) ou qu’il n’y a pas
(fig. 10.1 et 10.2, à gauche) un crossing-over entre B et C.
Remarque. Il n’y a pas d’autre scénario possible car les deux gènes A et B sont proches
de leurs centromères respectifs; il n’y a donc pas de crossing-over possible entre le
locus d’un gène et son centromère, ce qui conduirait une même méiose à produire
quatre types de gamètes différents, deux parentaux plus deux recombinés.
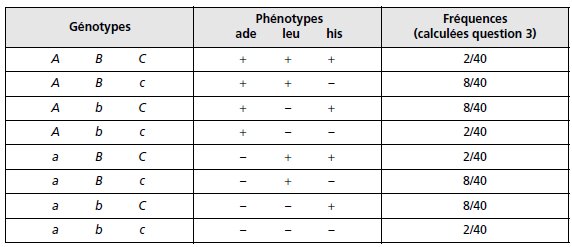
Les phénotypes des spores sont faciles à déduire de la présence ou de l’absence des mutations
d’auxotrophie (tabl. ci-dessous, colonnes 1 et 2), sachant qu’il existe huit combinaisons
possibles issues du brassage génétique pour trois gènes dialléliques (23 !).
ENSEMBLE DES GÉNOTYPES POSSIBLES DES SPORES
ISSUES DES MÉIOSES DÉCRITES FIGURES 10.1 ET 10.2.
Remarque. Une spore (gamète) comme (a; B; C) peut être parentale relativement à
deux gènes (ici le couple de gènes A et B) et recombinées relativement à deux autres
(ici le couple de gènes A et C ou le couple de gènes B et C).
Figure 10.1 Première disposition métaphasique possible (scénario 1)
de probabilité 1/2 sur l’ensemble des méioses.
λ figure la probabilité d’avoir un crossing-over entre B et C.
Figure 10.2 Deuxième disposition métaphasique possible (scénario 2)
de probabilité 1/2 sur l’ensemble des méioses.
λ figure la probabilité d’avoir un crossing-over entre B et C.
3. Si on désigne par λ la probabilité d’avoir un crossing-over entre les locus des gènes B et C,
alors les scénarios 1-a et 2-a ont, sur l’ensemble des méioses, une fréquence égale à (1 – λ),
soit une fréquence individuelle égale à (1 – λ)/2, puisque chacun d’entre eux est équifréquent
du fait de la disposition aléatoire des paires de chromatides non homologues, et la fréquence
des scénarios 1-b ou 2-b est égale à λ /2.
La distance entre les locus des gènes B et C est égale à 20 unités de recombinaison, ce qui
signifie que la fréquence des spores recombinées pour ces deux gènes est égale à 20 %.
Or ces spores recombinées, pour les gènes B et C, représentent la moitié des spores issues des
méioses des scénarios 1-b et 2-b, ce qui signifie que (λ/2 + λ/2)/2 = 0,20, d’où λ = 0,40.
Chacun des scénarios 1-a et 1-b a une fréquence égale à 0,30, et chacun des scénarios 1-b
et 2-b a une fréquence égale à 0,20.
Connaissant les fréquences des scénarios et les fréquences des différents types de spores qui
en résultent, il est facile de calculer les fréquences des huit types possibles de spores
(tabl. p. 269, colonne 3).
Remarque. Le fait que les locus des gènes A et B soient très proches de leurs centromères
respectifs limite le nombre de scénarios possibles de méioses.
En effet, si le locus du gène A était situé assez loin de son centromère pour qu’un
éventuel crossing over puisse survenir entre eux, on voit bien que chacun des quatre
scénarios se subdiviserait de nouveau en deux cas, avec ou sans crossing-over entre A
et son centromère. Cela ferait huit scénarios possibles, et même seize, si on admet
qu’un crossing-over puisse aussi survenir entre le locus du gène B et son centromère.
Cela ne changerait pas le nombre de types différents de spores, qui resterait toujours égal à
huit, mais cela changerait leurs fréquences respectives, en fonction des fréquences des différents
scénarios.
Il convient en effet de noter que les scénarios 1-a et 2-a qui conduisaient toujours, pour les
gènes A et B, à quatre gamètes parentaux, deux à deux identiques, ou à quatre gamètes
recombinés, deux à deux identiques, conduiraient, dans le cas d’un éventuel crossing-over
entre le locus du gène A et son centromère, à quatre gamètes tous différents, deux parentaux
et deux recombinés.
Ce phénomène est facile à mettre en évidence chez certains organismes (champignons ascomycètes)
où il est possible de faire l’analyse génétique des produits de chaque méiose isolément
, ce qui n’est jamais le cas dans la plupart des espèces où
l’analyse génétique est une analyse de gamètes en vrac, soit directe quand il s’agit de spores
qui peuvent donner des clones ou des organismes haploïdes (mousses, champignons), soit
indirecte, par des croisements, quand la phase haploïde est réduite aux gamètes (la plupart
des végétaux, et les animaux).