On analyse les tétrades obtenues après sporulation du diploïde issu du croisement
entre pho5 et une souche SR2 sauvage pour le phénotype phosphatase
acide, noté [pho+], et le phénotype tryptophane, noté [trp+], auxotrophe
[lys–] par mutation de perte de fonction dans le gène LYS2; les phénotypes
mutés ou sauvages sont respectivement notés [pho–], [trp–] et [lys+].
On observe les résultats suivants, classés par ordre d’importance, dont vous
donnerez une interprétation cartographique précise.
Question 3.
On analyse les tétrades obtenues après sporulation de diploïdes issus du
croisement entre pho5 et pho3, pour le phénotype phosphatase acide.
On
observe les résultats suivants, classés par ordre d’importance, dont vous
donnerez une interprétation cartographique précise.
Question 4.
On dispose d’un test colorimétrique qui permet de colorer en rouge les
colonies présentant une activité phosphatase acide supérieure à 7 unités
d’activité spécifique.
À partir d’une autre culture de la souche SR1 on isole, sur ce milieu test 10
mutants [pho–] donnant des colonies blanches et notés m1 à m10 (mutants
du problème de contrôle continu).
a. L’analyse génétique de ces 10 mutants révèle les faits suivants :
– les phénotypes [pho–] sont récessifs sauf deux, m3 et m7 qui sont
dominants;
– les mutants semblent mutés dans un seul gène (50 % de spores [pho+] et
50 % de spores [pho–] à l’issue de la méiose d’un diploïde formé par le
croisement entre chaque mutant mi et la souche sauvage SR2);
– l’analyse des croisements entre mutants récessifs permet de définir trois
groupes de complémentation (m1, m4, m9), (m2, m5, m8, m10) et (m4,
m6, m9);
– les différents croisements entre les mutants m1, m4, m6 et m9 permettent,
à l’issue de la méiose du diploïde, de recueillir quelques spores
sauvages [pho+], ce qui permet de faire la carte fine des mutations m1,
m4, m6 et m9, révélant qu’elles sont très liées et dans l’ordre suivant :
Quelle est la seule hypothèse génétique permettant de concilier ces observations,
sachant, par ailleurs que la colonie m6 est mutée dans un gène
PHO4 dont le produit forme des oligomères ?
b. Analyse biochimique du mutant PHO4 :
Figure 10.5
Sur un milieu carencé en Pi, mais avec une source organique de phosphate,
le mutant pousse très lentement.
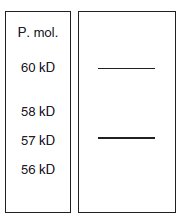
Le dosage, après culture sur milieu carencé en Pi, de l’activité spécifique et
la réalisation du profil électrophorétique conduit au résultat de la
figure 10.5, avec une activité spécifique autour de 7.
En présence de Pi, ce mutant pousse normalement.
L’analyse d’autres mutants du gène PHO4 a permis d’isoler un mutant
nommé PHO4c.
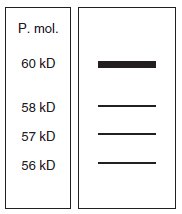
Le dosage, après culture sur milieu carencé en Pi, mais aussi sur milieu
riche en Pi, de l’activité spécifique et la réalisation du profil électrophorétique
conduit au résultat de la figure 10.6, avec une activité spécifique égale
à 30.
On obtient le même résultat pour un diploïde issu du croisement entre
PHO4c et une souche SR.
Après avoir tiré de l’analyse génétique et biochimique les informations
concernant les mutations pho4 et pho4c touchant respectivement les mutants
PHO4 et PHO4C, vous préciserez quel est l’effet de ces mutations et quelle
peut être la fonction du gène PHO4.
Vous justifierez votre réponse et vous
ferez un schéma précisant les interactions entre le gène PHO4 et les autres
gènes définis auparavant.
Question 5.
Sur des boîtes du milieu test qui permet la coloration en rouge des colonies
si l’activité phosphatase acide dépasse un seuil supérieur à 7 unités d’activité
spécifique, le mutant PHO4 donne des colonies blanches.
L’étalement
d’une culture mutagénisée de ce mutant PHO4 permet d’isoler quelques
colonies rouges.
L’une de ces colonies est croisée avec la souche SR2; à l’issue de la sporulation
du diploïde on observe quelques tétrades avec quatre spores [lys+].
Que pouvez vous en conclure ?
Question 6.
On a montré que le mutant m7 est muté dans un gène nommé PHO80,
physiquement indépendant de tous les autres gènes étudiés auparavant.
L’analyse biochimique de m7 donne des résultats identiques à ceux d’un
mutant PHO4 comme m6 (figure 2).
On rappelle cependant que, croisés
avec une souche SR2, les mutants m6 et m7 donnent des diploïdes de
phénotypes [pho+] pour l’un et [pho–] pour l’autre.
L’étalement d’une
culture après mutagenèse du mutant m7, sur des boîtes de milieu test colorimétrique,
permet d’isoler des colonies rouges dont l’analyse biochimique
se révèle identique à celle de la figure 3.
L’analyse génétique de ces colonies
permet de distinguer deux types de mutants, d’une part des mutants du
gène PHO80 lui-même dont l’analyse fonctionnelle permet de montrer
que ce sont des mutants de perte de fonction, d’autre part des mutants du
gène PHO4 qui se révèlent être du type PHO4c.
Compte tenu de toutes ces informations génétiques et biochimiques apportées
par l’étude du mutant PHO80, vous préciserez quel est l’effet de ces
mutations et quelle peut être la fonction du gène PHO80.
Vous justifierez
votre réponse et vous ferez un schéma précisant les interactions entre le gène
PHO80 et les autres gènes définis auparavant, notamment le gène PHO4.
➤ Niveau L3-Master :
Solution
1. Analyse génétique.
a. Les quatre mutants PHO3, PHO5, PHO10 et PHO11 sont récessifs puisque les diploïdes
issus de leurs croisements respectifs avec une souche sauvage SR1 [pho+] sont de phénotype
sauvage.
b. Ils semblent mutés dans un seul des gènes impliqués dans l’activité phosphatase acide,
puisqu’on observe une ségrégation 2/2 des phénotypes [pho+] et [pho–] dans la méiose des
diploïdes mutant × SR1.
Leurs mutations seront respectivement désignées par pho3, pho5,
pho10 et pho11.
c. Les croisements entre mutants récessifs peuvent être interprétés sur le plan fonctionnel
comme un test de complémentation ou de non complémentation indiquant si les mutants sont
mutés dans un (le) même gène ou non.
On peut conclure, à partir des résultats, que les quatre
mutants sont mutés dans des gènes différents.
2. Analyse biochimique qualitative et quantitative.
a. Elle confirme le résultat de l’analyse génétique, à savoir que les quatre mutants ne semblent
mutés que dans un seul gène, différent pour chacun :
– la mutation pho3 est une perte de fonction dans le gène codant une phosphatase alcaline de
57kD,
– la mutation pho5 est une perte de fonction dans le gène codant une phosphatase alcaline de
60kD,
– la mutation pho10 est une perte de fonction dans le gène codant une phosphatase alcaline
de 58kD,
– la mutation pho11 est une perte de fonction dans le gène codant une phosphatase alcaline
de 56kD.
b. Le gène PHO5 est plus exprimé et/ou son produit est plus actif puisque l’activité résultante
qui lui est attachée est plus forte.
c. Les trois gènes PHO5, PHO10 et PHO11, ont une expression régulée puisqu’ils sont
exprimés en absence de Pi mais ne sont plus exprimés en présence de Pi, soit qu’ils sont
réprimés, soit qu’ils ne sont pas activés.
Le gène PHO3 est exprimé de façon constitutive puisqu’il n’est pas régulé par la présence ou
l’absence de Pi.
d. Le gène PHO5, bien que régulé, laisse apparaître la présence d’une quantité faible d’activité
même en condition de répression ou de non activation.
Cette quantité minimale, ou taux
de base, semble indiquer que la quantité importante de produit PHO5 correspond à une forte
expression du gène PHO5 plutôt qu’à une forte activité de son produit, de sorte qu’il demeure
une transcription minimale en condition de répression ou de non activation.
2. L’analyse les tétrades issues de la méiose d’un diploïde issu de l’union de spores [pho+, trp+]
et [pho–, trp–] permet de définir les spores parentales et recombinées. On identifie 24 ditypes
parentaux (DP), dont les quatre spores sont parentales, 22 ditypes recombinés (DR), dont les
quatre spores sont recombinées, et 4 tétratypes (T).
L’égalité de fréquence entre DP et DR permet de conclure à l’indépendance génétique entre
les locus des gènes PHO5 et TRP1.
En outre, la fréquence des T étant significativement inférieure
à 2/3, on peut conclure à leur indépendance physique.
Enfin, le gène TRP1 étant un marqueur centromérique, on peut conclure que les T ne peuvent
être dus qu’à une post-réduction entre le locus du gène PHO5 et son centromère, ce qui
permet d’estimer la distance de PHO5 à son centromère, la fréquence des tétratypes étant
alors égale à la fréquence de post-réduction pour PHO5.
La distance est égale à la demifréquence
de post-réduction, soit la demi-fréquence des tétratypes, soit 0,04 × 100 = 4 unités
de distance centromérique par post-réduction.
3. Le croisement entre mutants pho5 et pho3 concerne des gènes PHO5 et PHO3 différents
(il y a complémentation fonctionnelle, voir question 1).
Les spores [pho+] observées dans certaines tétrades résultent donc de la recombinaison entre
ces deux gènes conduisant à deux spores, l’une sauvage et l’autre double mutante.
On attend
donc, pour des DR, la présence de deux spores [pho+] et de deux spores [pho–].
L’analyse de
tétrades montre qu’on obtient 80 DP, 19 T et 1 DR.
Cette observation permet de conclure que les gènes PHO5 et PHO3 sont génétiquement (et
physiquement) liés puisque les DP (donc les spores parentales) sont significativement plus
nombreux que les DR (donc les spores recombinées).
Elle permet aussi d’estimer une distance génétique entre les deux sites (gènes) mutés,
distance corrigée par l’analyse de tétrades, soit 100[f(T)/2 + 3f(DR)] = 12,5 unités de chromatides
remaniées (ou cM si on estime que les distances corrigées sont additives).
4. Analyse génétique.
L’analyse de la complémentation fonctionnelle permet de conclure d’une part que les mutants
m1, m4 et m9 sont mutés dans un même gène, d’autre part que les mutants m4, m6 et m9
sont aussi mutés dans un même gène, ce qui pourrait laisser croire que les mutants m4 et m9
sont au moins des doubles mutants.
Or une autre analyse (croisement par sauvage) semble montrer que tous ces mutants sont
simples, mutés dans un seul gène, ce qui serait contradictoire pour m4 et m9.
Et la carte fine, compte tenu de la position du site m1, est également cohérente avec l’hypothèse
que m1, m4, m6 et m9 sont chacun mutés dans un seul et même gène.
On pourrait alors
imaginer que les mutants m4 et m9 soient des mutants par délétion (d’où la ségrégation 2-2
chez les diploïdes avec SSR) touchant deux gènes contigus, mais alors elles devraient toutes
deux couvrir le site m1 ce qui est contradictoire avec le fait qu’on a des recombinants sauvage
[pho+].
Il faut donc admettre que les quatre sites mutés appartiennent au même gène et que la complémentation
fonctionnelle chez les diploïdes m1 × m6 serait de la complémentation intragénique.
Cette hypothèse est cohérente avec le fait que le gène concerné, PHO4, celui qui est muté
chez m6 et donc aussi les autres mutants m1, m4 et m9, code pour une chaîne peptidique
formant des homo-dimères.
On sait en effet qu’une mutation de perte de fonction touchant un
site d’interaction sur un protomère peut voir son effet corrigé ou supprimé par une autre
mutation touchant le même site sur un autre protomère et permettant alors l’interaction entre
les deux protomères mutés chez un diploïde porteur des deux mutations différentes.
Les mutants m1 et m6 présentent donc une perte de fonction liée à une mutation dans un site
d’interaction entre protomères, alors que les mutants m4 et m9 présentent une perte de fonction
dans un domaine actif du protomère, type de perte de fonction qui ne permet pas la complémentation intragénique.
Analyse biochimique du mutant PHO4.
Le dosage d’activité chez PHO4 est effondré à 7, et on ne distingue que l’activité PHO3, qui est
constitutive, et l’activité PHO5, qui est à son taux de base, ce qui permet de conclure qu’aucun
des gènes régulés n’est exprimé.
Or il s’agit d’un mutant simple, muté dans un seul gène, ce qui conduit à l’hypothèse que le
gène PHO4 serait un gène de régulation.
– si PHO4 est un gène codant pour un activateur, tous les mutants de phénotype [pho–],
comme m1, m4, m6 ou m9, seraient porteurs d’une mutation pho4 de perte de fonction,
dont l’effet serait récessif vis-à-vis de celui de l’allèle sauvage pho4+,
– si PHO4 est un gène codant pour un répresseur, les mutants de phénotype [pho–] seraient
alors porteurs d’une mutation de gain de fonction, notée pho4+++, rendant le répresseur
super-actif, voire actif même en présence de Pi, par exemple par incapacité de se lier au
ligand-inducteur chargé de le rendre inactif dans les conditions où les gènes sous sa dépendance
doivent être transcrits (c’est-à-dire en absence de Pi). Mais, dans ce cas, on s’attendrait
à ce que le phénotype [pho–] soit dominant, ce qui n’est pas le cas.
D’ailleurs l’observation du mutant PHO4c chez lequel l’expression de tous les gènes est
constitutive (expression sauvage à la valeur 30, même en milieu riche en Pi où les activités
phosphatases ne sont pas requises) est cohérente avec l’hypothèse que PHO4 code pour un
activateur, que les mutants de phénotype [pho–] sont des mutants de perte de fonction (pho4)
et que le mutant PHO4c serait touché dans le site de liaison avec le ligand-inhibiteur. Dans ce
cas le phénotype constitutif doit être dominant, ce qui est le cas.
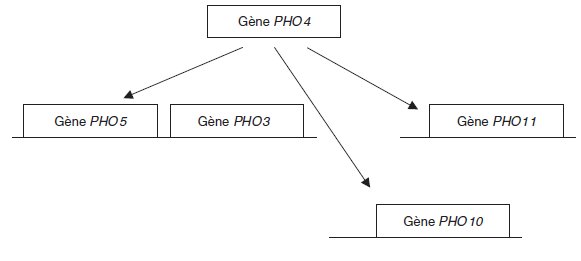
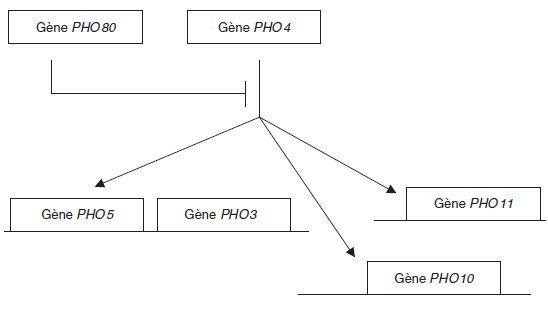
D’où le schéma précédent, où les flèches indiquent que le produit PPHO4 du gène PHO4 est
un activateur des gènes PHO5, PHO10 et PHO11, le gène PHO3 étant constitutif.
5. Les colonies rouges obtenues après mutagenèse d’un mutant pho4 ont un phénotype sauvage
[pho+] et sont des révertants, éventuellement porteurs d’une mutation suppresseur capable de
supprimer l’effet de la mutation directe de perte de fonction touchant le mutant pho4.
Pour le phénotype lysine, conditionné par la situation génétique au locus du gène LYS2, le
diploïde est lys2+/lys2– et doit obligatoirement donner deux spores de génotype (lys2+) et
deux spores de génotype (lys2–), soit deux spores [lys+] et deux spores [lys–], à moins que le
révertant [pho+] soit un révertant par suppresseur informationnel capable de corriger à la fois
l’effet de la mutation pho4 et celui de la mutation lys2, ce qui est ici le cas, puisque certaines
tétrades présentent quatre spores [lys+].
6. Le mutant m7 est muté dans le gène PHO80, mais il s’agit d’un mutant dominant, contrairement
à m6, muté dans PHO4.
Son analyse biochimique révèle aussi l’absence de toute expression des trois gènes habituellement
soumis à une régulation (avec l’expression de PHO5 à son taux de base).
On pourrait, a priori, faire ici la même interprétation que pour PHO4 (question 4/b) à savoir
que PHO80 code soit pour un activateur, soit pour un répresseur; mais il serait difficile
d’interpréter le fait que le mutant m7 soit dominant, car une perte de fonction dans un activateur
est rétablie chez le diploïde avec SR et la dominance de m7 oriente plutôt l’interprétation
vers l’hypothèse d’un mutant dans un répresseur touché dans son site de liaison avec un
ligand.
Les révertants sont de deux types :
– soit des mutants de perte de fonction dans le gène PHO80 lui-même, ce qui est cohérent
avec l’hypothèse que ce gène coderait pour un répresseur, et que le mutant m7 serait un
mutant dans le site de liaison du ligand rendant le répresseur super-actif;
– soit des mutants du type PHO4c, chez lequel il faut considérer que le répresseur codé par le
gène PHO80 est toujours présent mais inactif puisque le phénotype du révertant est [pho+],
ce qui ne peut s’expliquer qu’en supposant que le répresseur codé par PHO80 n’est pas un
répresseur de la transcription agissant sur les gènes PHO5, PHO10, PHO11 ou même PHO4
mais qu’il agit directement sur l’activateur produit par le gène PHO4, c’est-à-dire la protéine
PPHO4.
Les mutants PHO4c sont alors insensibles à l’action du répresseur codé par PHO80.
Ce résultat illustre l’importance de la recherche et de l’analyse génétique et biochimique des
révertants dans l’analyse d’un système biologique.
On peut alors proposer le schéma précédent, où le segment barré issu du gène PHO80 illustre
le fait que son produit est un répresseur qui bloque l’action de l’activateur produit par le gène
PHO4.