1. Le fait d’avoir des colonies sur milieu minimum prouve que les trois phénotypes d’auxotrophie
sont récessifs et que les souches A et B sont mutées dans, au moins, trois gènes différents
de sorte qu’il y a complémentation fonctionnelle chez le diploïde qui est le seul à
pouvoir se développer sur la boîte d’étalement.
La question se pose alors de savoir si chaque phénotype d’auxotrophie ne dépend que d’un
seul gène.
2. • Nombre de gènes en jeu pour chaque phénotype d’auxotrophie
La question de savoir si chaque auxotrophie ne dépend que d’un seul gène est résolue par le
test de ségrégation 2/2 à la méiose du diploïde.
Dans les cinquante tétrades on décompte, pour chaque phénotype, met ou trp ou his,
100 spores sauvages [+] et 100 spores mutées auxotrophes [–], ce qui est le résultat attendu
d’une ségrégation 2/2 d’un couple d’allèles.
Chaque phénotype d’auxotrophie ne dépend que
d’un seul gène, A est mutée dans un des gènes de la chaîne de biosynthèse de la méthionine
et dans un des gènes de la chaîne de biosynthèse de l’histidine, et B est mutée dans un des
gènes de la chaîne de biosynthèse du tryptophane.
• Indépendance ou liaison des gènes
L’analyse de tétrades pour les deux gènes impliqués dans les auxotrophies met et trp permet
d’observer 24 DP et 26 DR.
Ces deux gènes sont génétiquement indépendants.
La fréquence des tétratypes étant nulle, donc inférieure à 2/3, on peut conclure que ces deux
gènes sont non seulement physiquement indépendants, mais également très proches de leurs
centromères respectifs.
L’analyse de tétrades pour les deux gènes impliqués dans les auxotrophies met et his permet
d’observer 18 DP et 20 DR.
Ces deux gènes sont génétiquement indépendants.
La fréquence des tétratypes étant inférieure à 2/3, on peut conclure que ces deux gènes sont
physiquement indépendants.
Comme les tétratypes, pour deux gènes physiquement indépendants, résultent de la survenue
d’au moins un crossing entre un gène et son centromère, et que cet événement est impossible (ou excessivement rare) pour le gène impliqué dans l’auxotrophie met, on peut conclure que
les tétratypes résultent tous d’une postréduction pour le gène impliqué dans l’auxotrophie
his, ce qui permet alors d’identifier la fréquence des tétratypes à la fréquence de postréduction
pour ce gène, et de calculer ainsi sa distance au centromère, soit 12 upr (la fréquence de
postréduction étant égale à 12/50 = 0,24).
Remarque 1. L’estimation de la postréduction et le calcul de la distance au centromère
peuvent être réalisés directement dans l’étude de tétrades ordonnées.
Chez la levure où les tétrades ne sont pas ordonnées, l’utilisation de marqueurs centromériques, des gènes très liés à leurs centromères, comme ici ceux impliqués
dans les auxotrophies met ou trp, permet d’estimer indirectement la fréquence de
postréduction d’un gène par l’estimation de la fréquence de tétratypes pour ce gène et
le marqueur centromérique.
Remarque 2. Le nombre de tétratypes pour les deux auxotrophies trp et his est obligatoirement
le même que pour les deux auxotrophies met et his.
En effet les deux
gènes « met » et « trp » ségrégeant avec leurs centromères respectifs, un tétratype ne
peut survenir qu’à l’issue d’un crossing-over entre le gène « his » et son centromère
et, dans ce cas, il s’agit aussi bien d’un tétratype relativement à his et met que relativement
à his et trp.
3. Si 50 spores sont de phénotypes [gal–] il ne peut pas s’agir de l’apparition d’un mutant
spontané, cela signifie qu’une des deux souches A ou B était mutée dans un (ou plusieurs)
gène(s) de métabolisation du galactose, mais était aussi porteuse d’une mutation ayant un
effet suppresseur.
Autrement dit, l’une des deux souches est un « révertant » [gal+], de génotype (gal–; sua),
issu d’un mutant [gal–], dont le génotype était (gal–; sui), où gal– correspond à la mutation
directe (en supposant qu’il s’agit d’un mutant simple, dans un seul gène) et sua correspond à
la mutation suppresseur actif (sui étant la séquence sauvage).
Le croisement A × B correspond donc, pour le phénotype gal, à un croisement révertant
× sauvage et permet, à la méiose du diploïde (gal–/gal+; sua/sui), de mettre en évidence l’existence
du suppresseur par la recombinaison génétique qui peut le séparer de la mutation
directe et faire ainsi apparaître des spores recombinantes mutées (gal–; sui) de phénotype [gal–].
Les 40 tétrades avec 4 spores [gal+] sont obligatoirement des DP; les DR ayant obligatoirement
au moins deux spores recombinantes [gal–] sont au nombre de 1, et les 9 autres sont
donc des T, ce qui permet de conclure que :
– la spore de génotype (gal–; sua) est de phénotype [gal+];
– la mutation gal– est génétiquement et physiquement liée à la mutation sua ;
– la distance corrigée entre ces deux mutations est égale à 15 ur, soit 100 × (f [T]/2 + 3f [DR]).
Problème 10.6
On admettra, dans tout le problème qu’on dispose, pour les croisements,
de spores de signe sexuel adéquat avec les marqueurs de sélection
nécessaires.
On dispose de deux souches haploïdes de levure Saccharomyces cerevisiae,
une souche SSR et une souche A, auxotrophe pour le tryptophane, de phénotype [trp–], porteuse dans le gène TRP1 de la mutation opale trp1-5.
Le locus du gène TRP1 est contigu au centromère.
À partir de la souche A, on obtient une collection de mutants auxotrophes
pour l’arginine et de phénotype [Arg–; Trp–].
On s’intéresse à des mutants simples, mutés soit dans le gène ARG1 soit
dans le gène ARG2, dans le but d’analyser les rapports cartographiques et
fonctionnels entre certains mutants de ces deux groupes.
1. Analyse de mutants dans ARG1 : arg1-1, arg1-2 et arg1-3
a. Le mutant arg1-1, de phénotype [arg–; trp–] est croisé avec la souche
SSR, puis mis à sporuler afin d’étudier les tétrades obtenues.
On a :
– 12 tétrades avec deux spores [arg–; trp–] et deux spores [arg+; trp+];
– 10 tétrades avec deux spores [arg–; trp+] et deux spores [arg+; trp–];
– 13 tétrades avec une spore [arg–; trp–], une spore [arg+; trp+], une spore
[arg–; trp+] et une spore [arg+; trp–].
Quelles sont toutes les conclusions que ce résultat permet d’établir ?
b. On entreprend cinq expérimentations.
• Expérimentation 1.
Les diploïdes arg1-1 × arg1-2; arg1-1 × arg1-3 et
arg1-2 × arg1-3 sont mis à sporuler. On recueille un grand nombre de
spores de chacun de ces croisements afin de tester, à chaque fois, environ
100 000 spores en les déposant sur une boîte de milieu Mo additionné de
tryptophane.
Avec les spores issues du premier croisement, on obtient 75 colonies; avec
celles issues du deuxième, on obtient 50 colonies; avec celles issues du
troisième, on n’en obtient aucune, même en répétant l’expérience.
• Expérimentation 2.
Des cultures de chacun des trois mutants arg1-1,
arg1-2 et arg1-3, soumises à différents types de mutagènes, sont étalées
sur un milieu Mo additionné de tryptophane. Dans les deux premiers cas,
on obtient quelques colonies, jamais dans le troisième.
• Expérimentation 3.
Les colonies obtenues précédemment sur milieu Mo
additionné de tryptophane, et issues de la mutagenèse de mutants arg1-2,
sont repiquées sur Mo.
La plupart d’entre elles se révèlent capables d’y
pousser.
• Expérimentation 4.
Les colonies obtenues sur milieu Mo additionné de
tryptophane, et issues de la mutagenèse de mutants arg1-1, sont repiquées
sur Mo. Elles sont toujours incapables d’y pousser.
• Expérimentation 5.
Les colonies obtenues sur milieu Mo additionné de
tryptophane, et issues de la mutagenèse de mutants arg1-1, avec des agents intercalants, sont repiquées sur Mo.
Elles sont toujours incapables d’y
pousser.
En vous appuyant sur ces cinq expérimentations et en justifiant avec rigueur
vos réponses :
– vous donnerez une interprétation cartographique précise (avec schéma)
de ces résultats;
– vous préciserez, autant qu’il est possible, le type de mutation (non-sens,
faux-sens, décalage, délétion…) affectant chacun des mutants étudiés
arg1-1, arg1-2 et arg1-3.
2. Analyse d’un mutant dans ARG2 : arg2-1
Le mutant arg2-1, de phénotype [arg–; trp–] est croisé avec la souche SSR,
puis mis à sporuler afin d’étudier les tétrades obtenues.
On observe les
résultats suivants, à interpréter :
– 87 tétrades avec deux spores [arg–; trp–] et deux spores [arg+; trp+];
– 1 tétrade avec deux spores [arg–; trp+] et deux spores [arg+; trp–];
– 12 tétrades avec une spore [arg–; trp–], une spore [arg+; trp+], une spore
[arg–; trp+] et une spore [arg+; trp–].
3. On extrait l’ADN du mutant arg1-1 et on le soumet à une digestion
partielle avec l’endonucléase Sau3A afin d’insérer les fragments obtenus
dans un plasmide réplicatif multicopies, porteur de l’allèle sauvage trp1+
du gène TRP1.
On transforme, avec les plasmides recombinants ainsi obtenus, la souche
arg1-1 et on étale sur un milieu Mo additionné d’arginine.
On obtient
environ 20 000 colonies qui sont repiquées sur Mo; il y pousse 5 colonies.
Deux hypothèses peuvent permettre d’expliquer ce résultat, lesquelles ?
Puis on extrait les plasmides de chacune des 5 colonies précédentes afin de
retransformer la même souche arg1-1.
Dans 1 cas, la souche retransformée
est de phénotype [trp+; arg–], dans 4 cas, elle est de phénotype [trp+; arg+].
Précisez le but de cette expérimentation et les précisions qu’elle apporte.
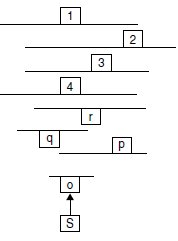
4. Les 4 plasmides conférant, par transformation, le phénotype [trp+; arg+]
à la souche arg1-1 sont étudiés in vitro; les 4 inserts présentent des cartes
de restriction se recouvrant en partie (fig. 10.3).
a. Qu’en concluez-vous ?
b. Par digestion avec des endonucléases, on isole les sous-fragments o, p, q
et r, tels qu’indiqués ci-dessus. Ces fragments sont clonés dans un plasmide
réplicatif multicopie porteur de l’allèle sauvage trp1+ de TRP1.
On transforme, avec chacun de ces plasmides, la souche (arg1-1; trp1-5) :
– tous les transformants de phénotype [trp+] sont de phénotype [arg–],
quand le plasmide utilisé est porteur des fragments o, p ou q;
– tous les transformants de phénotype [trp+] sont de phénotype [arg+],
quand le plasmide utilisé est porteur du fragment r.
Qu’en concluez-vous ?
Figure 10.3
1, 2, 3 et 4 : inserts des plasmides conférant le phénotype [trp+, arg+] à
arg1-1; o, p, q et r : sous-fragments des inserts testés pour leur capacité à
restaurer le phénotype [arg+]; S : site Sal1, interne au fragment o.
c. Le fragment O est cloné dans un plasmide intégratif, porteur de l’allèle
sauvage trp1+ de TRP1; celui-ci est alors linéarisé au site S (Sal1) et utilisé
pour transformer la souche A :
– tous les transformants sont [trp+; arg–];
– ces transformants [trp+; arg–] croisés avec une SSR donnent des
diploïdes [arg+];
– ces transformants, croisés avec le mutant arg2-1, donnent des diploïdes
de phénotype [trp+; arg–].
Qu’en concluez-vous ?
d. L’un de ces transformants de phénotype [trp+; arg–] est croisé avec la
souche SSR afin d’étudier les phénotypes des spores issues de la méiose.
On observe les résultats suivants :
– spores [trp+, arg+] 45,5 %;
– spores [trp+, arg–] 50 %;
– spores [trp–, arg+] 4,5 %;
– spores [trp–, arg–] 0 %.
En quoi ce résultat confirme-t-il la conclusion de la question précédente ?
5. Finalement, quelle hypothèse pourriez-vous faire sur la nature de la
mutation arg1-1, le mode d’action de son suppresseur et la relation fonctionnelle
entre les gènes ARG1 et ARG2, sachant qu’aucune mutation du
gène ARG1 n’a été trouvée plus en 5′ que la mutation arg1-1 ?