Remarque. À ce stade du problème, cette dernière hypothèse n’est pas attendue parmi
les réponses des étudiants, même si l’énoncé de la question précise que le génotype de
A est « présumé ».
4. Dans ce croisement, seuls les phénotypes tryptophane et lysine sont analysés, ce qui
concerne dans chaque croisement trois gènes, les gènes TRP1, LYS2 et le gène suppresseur,
nommé su1 dans RA1, su2 dans RA2 et su3 dans RA3.
Dans chaque croisement le diploïde obtenu est de génotype :
où :
– le premier allèle est celui apporté par le révertant et le second allèle est celui apporté par la
souche (met14-5, his2-3);
– n est le numéro du suppresseur (1, 2 ou 3), dans sa version allélique active (a) ou inactive,
sauvage (i);
– les pointillés indiquent l’ignorance relative à une liaison génétique éventuelle du locus du
suppresseur avec l’un des deux locus précédents (ceux-ci étant physiquement indépendants,
voir énoncé).
À partir de ce génotype, et en considérant le gène suppresseur et le gène TRP1, quatre types
de spores sont possibles :
– spore parentale : (trp1-3, sun
a) de phénotype [trp+] pour n = 1, 2 ou 3,
– spore parentale : (trp1+, sun
i) de phénotype [trp+] pour n = 1, 2 ou 3,
– spore recombinée : (trp1-3, sun
i) de phénotype [trp–] pour n = 1, 2 ou 3,
– spore recombinée : (trp1+, sun
a) de phénotype [trp?] pour n = 1, 2 ou 3.
Si la spore recombinée (trp1+, sun
a) avait un phénotype [trp–], on observait parmi les trois
types de tétrades, des DR avec quatre spores [trp–], ce qui n’est pas le cas et montre que le
phénotype est [trp+]
À partir de ce génotype, et en considérant le gène suppresseur et le gène LYS2, quatre types
de spores sont possibles :
– spore parentale : (lys2-4, sun
a) de phénotype [lys+] pour n = 1, 2 mais pas pour n = 3,
– spore parentale : (lys2+, sun
i) de phénotype [lys+] pour n = 1, 2 ou 3,
– spore recombinée : (lys2-4, sun
i) de phénotype [lys–] pour n = 1, 2 ou 3,
– spore recombinée : (lys2+, sun
a) de phénotype [lys?] pour n = 1, 2 ou 3.
Si la spore recombinée (lys2+, sun
a) avait un phénotype [lys–], on observait parmi les trois
types de tétrades, des DR avec quatre spores [lys–], ce qui n’est pas le cas et montre que le
phénotype est [lys+].
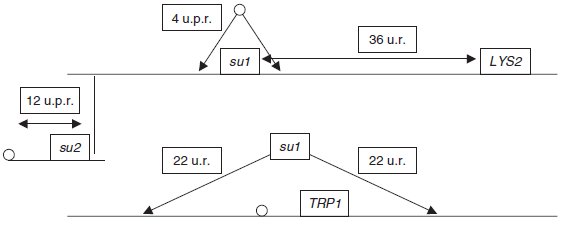
L’analyse du tableau conduit alors aux conclusions suivantes :
a. Le suppresseur su1
a de RA1 est :
– génétiquement indépendant de TRP1 (22 DR et 24 DP) et physiquement également (4 T, très
inférieur à 2/3),
– génétiquement lié à LYS2 (34 DP, 4 DR), ce qui conduit à une distance génétique corrigée
égale à 100(12/2 + 3 × 4)/50 = 36 u.r.,
– à une distance calculable de son centromère, car TRP1 étant centromérique, tous les tétratypes
résultent d’une post-réduction entre le locus du suppresseur et son centromère; cette
distance est égale à 100(4/50)/2 = 4 u.p.r.
b. Le suppresseur su2
a de RA2 est :
– génétiquement indépendant de TRP1 (18 DR et 20 DP) et physiquement également (12 T,
très inférieur à 2/3),
– génétiquement indépendant LYS2 (9 DP, 8 DR), sans qu’on puisse statuer sur l’indépendance
physique puisque la fréquence des tétratypes est égale à 2/3,
– à une distance calculable de son centromère, car TRP1 étant centromérique, tous les tétratypes
résultent d’une post-réduction entre le locus du suppresseur et son centromère; cette
distance est égale à 100(12/50)/2 = 12 u.p.r.
c. Le suppresseur su3
a de RA3 est :
– génétiquement lié à TRP1 (38 DR et 2 Dr), ce qui conduit à une distance génétique
corrigée égale à 100(10/2 + 3 × 2)/50 = 22 u.r., qui constitue aussi la distance du suppresseur
à son centromère (voir remarque);
– génétiquement et physiquement indépendant LYS2, non par l’analyse de tétrades qui réunissent
toujours deux spores [lys+] et deux spores [lys–], qu’elles soient DP, DR ou T, mais parce
que su3 est lié à TRP1 qui, lui, est physiquement indépendant de LYS2 (voir énoncé).
Remarque.
Si on calculait la distance de su3 à son centromère en considérant TRP1
comme marqueur centromérique, elle serait égale à 100(10/50)/2 = 10 u.p.r. et serait sous
estimée, car on ne prendrait en compte, dans ce calcul, que la post-recombinaison
issue d’un seul crossing-over.
On peut présenter la cartographie suivante :
5.a. On a sept révertants où la chaîne peptidique du gène de structure ADA1 apparaît comme
sauvage; comme il est difficile d’imaginer qu’on ait sept révertants vrais, il est logique de
considérer que le mutant ada1-1 n’est pas muté dans le gène ADA1 mais dans un gène de
régulation de ce gène de structure, par exemple dans le gène d’un activateur par une mutation
de perte de fonction, ou dans le gène d’un répresseur, par un gain de fonction conduisant à un
répresseur super actif.
Remarque Pour pouvoir toucher le gène ADA1, on est parti de mutants (ade10-1,
dea2-1), capables de pousser sur hypoxanthine et adénine, pour obtenir des mutants
ayant perdu la capacité de pousser sur adénine. de tels mutants peuvent évidemment
être mutés dans ADA1 mais peuvent aussi être mutés dans un autre gène, comme c’est
ici le cas, ce qui montre bien que toute relation entre un phénotype et sa causalité génétique
suppose une analyse fine !…et qu’un même phénotype peut recouvrir des réalités
génétiques très différentes !
5.b. Si le révertant présente une chaîne peptidique active mais différente de celle du sauvage,
c’est que le mutant est bien touché dans le gène de structure concerné (s’il était touché dans
un gène de régulation de ce gène de structure, la chaîne peptidique du révertant serait sauvage,
comme dans le cas précédent).
Ici, on peut conclure que la mutation ada1-2 est une mutation
de décalage du cadre de lecture dans la séquence codante du gène ADA1, et que le révertant
présente une mutation de recalage (décalage en sens inverse) légèrement en amont ou en aval
du site de la mutation directe, de sorte que quelques acides aminés contigus sont modifiés
dans la partie du cadre qui est demeurée décalée entre les deux sites, sans que cette modification
altère la fonction de la chaîne peptidique.
5.c. Si la mutation nommée ada1-1 touchait le gène ADA1 et était alors allélique de la mutation
ada1-2, le génotype du diploïde serait ada1-1 // ada1-2.
Étant par ailleurs homozygote
ade10-1 // ade10-1 et dea2-1 // dea2-1, ce diploïde n’aurait pas la capacité de pousser sur
adénine (pas de fonction désaminase), ce qui est le cas.
Mais, si les mutations ada1-1 et ada1-2 étaient alléliques, l’analyse de tétrades ne devrait
alors donner que des spores [ade-] incapables de pousser sur adénine, ces spores étant toutes
déficientes pour les fonctions ADA1 (ségrégation 2-2 des allèles ada1-1 et ada1-2), DEA2 et
ADE10.
Or ce n’est pas le cas, ce qui confirme que ada1-1 et ada1-2 ne peuvent être alléliques.
Comme on doit considérer (voir question a) que la mutation ada1-1 ne touche pas le gène
ADA1 mais un autre gène, nommé R, le génotype du diploïde peut s’écrire :
R(ada-1-1) //R+ ----- ada1+//ada1-2
où :
– le premier allèle correspond à l’apport de la souche A et le second à l’apport de la souche B;
– l’allèle muté R correspond à l’allèle muté chez le mutant ada1-1;
– les pointillés indique l’ignorance quant à la liaison entre les locus des deux gènes.
Comme ce diploïde s’avère incapable de pousser sur adénine, on doit considérer que le
phénotype [ade-] sur milieu Mo avec adénine est dominant, que l’effet de la mutation R(ada1-1)
est dominant sur celui de l’allèle sauvage R+.
L’analyse de tétrades fait apparaître trois types de tétrades, ce qui est une nouvelle confirmation
de l’absence de ségrégation 2/2 pour R(ada1-1) et ada1-2.
La méiose peut conduire à quatre types de spores :
– spore parentale : (R, ada+) de phénotype [ade–],
– spore parentale : (R+, ada1-2) de phénotype [ade–],
– spore recombinée : (R+, ada+) de phénotype [ade+],
– spore recombinée : (R, ada1-2) de phénotype [ade–] si on considère l’effet joint des
deux mutations car l’analyse de tétrades donnerait
des tétrades DR à quatre spores [ade+] si le phénotype
n’était pas [ade–].
On a donc 17 DP et 18 DR, ce qui permet de conclure à l’indépendance génétique de ces
deux gènes, et seulement 12 tétratypes (moins de 2/3) ce qui permet de conclure à leur indépendance
physique.
La mutation ada1-1 n’affecte donc pas le gène ADA1 mais un autre gène R, physiquement
indépendant de ADA1 et spécifiant vraisemblablement un répresseur de ADA1, de sorte que
la mutation R (notée initialement ada1-1) serait un gain de fonction conduisant à la présence
d’un répresseur sur-actif, dont l’effet serait dominant chez le diploïde.
Il serait logique de
renommer la mutation ada1-1 en la désignant par R+++.
Chez les sept révertants RA4 à RA10, le phénotype [ade+] est recouvré par une mutation
suppresseur de perte de fonction affectant le gène R et abolissant l’effet du répresseur suractif
codé par l’allèle R+++.
Problème 10.9
On suppose, dans tout le problème, qu’on dispose des souches de signe
sexuel, et des marqueurs de sélection des diploïdes adéquats.
La cellule de levure S. cerevisiae a besoin d’une grande quantité de phosphate
pour sa croissance et sa multiplication (synthèse de ses acides
nucléiques et des phospholipides).
Les sources de phosphate qu’elle utilise
sont préférentiellement le phosphate inorganique libre externe (Pi) transporté
tel quel dans la cellule par une perméase, ou le phosphate (Pi) libéré
par clivage de composés organiques par des enzymes appelées phosphatases.
Des phosphatases acides (pH3 ou 4 optimum), membranaires ou périplasmiques
permettent l’utilisation de composés d’origine extérieure, la levure,
comme tous les champignons, pouvant se développer en milieu très acide.
Des phosphatases alcalines cytoplasmiques (pH8 optimum) permettent le
recyclage du phosphate à partir de métabolites phosphorylés.
On peut réaliser le dosage de l’activité phosphatase acide par une mesure
d’activité spécifique (activité rapportée à 1 mg de protéine) et on peut
réaliser la séparation électrophorétique de phosphatases de masse moléculaire
et/ou de charge différentes (figure 1), qui sont mises en évidence par
révélation spécifique de l’activité phosphatase acide dans le gel d’électrophorèse.
On a observé, chez S. cerevisiae, que l’activité enzymatique de type phosphatase
acide est très élevée en conditions de carence en Pi inorganique
dans le milieu extérieur, et très faible en présence de fortes concentrations
dans le milieu.
Ainsi la souche de levure haploïde de référence SR1 a une
activité de phosphatase de 30 unités d’activité spécifique en conditions de
carence en Pi externe et de 2 unités d’activité spécifique, en présence de fortes concentrations en Pi externe (voir figure 1).
La souche SR1 est de
génotype (his1–, trp1–), deux mutations d’auxotrophie responsables de
phénotypes récessifs [his–] et [trp–]; le gène TRP1, marqueur de centromère,
est situé assez près de son centromère pour que ses allèles soient
toujours pré-réduits.
On se propose au vu des résultats biochimiques, d’entreprendre
l’analyse des gènes impliqués dans cette activité phosphatase.
Question 1.
Dans des travaux précédents, on a pu isoler à partir de la souche SR1
quatre mutants, appelés PHO3, PHO5, PHO10 et PHO11, présentant une
activité phosphatase acide réduite. Le phénotype muté est caractérisé à la
fois par un dosage d’activité spécifique et par un profil électrophorétique
de cette activité (voir plus haut et figure 10.4).
L’analyse génétique et biochimique de ces mutants a conduit aux résultats
suivants :
– les diploïdes issus du croisement entre chaque mutant et une souche
sauvage pour le phénotype phosphatase acide, sont de phénotype sauvage
équivalent à celui de SR1 (voir figure 10.4);
– la sporulation de ces diploïdes conduit à deux types rigoureusement équifréquents
de spores de phénotype sauvage (du type SR1, voir figure 1)
ou de phénotype muté (voir figure 10.4);
– les croisements entre mutants donnent des diploïdes de phénotype
sauvage.
Proposez, avec concision, aussi bien pour les mutants que pour la souche
SR1, une interprétation exhaustive de ces résultats sur le plan génétique et
biochimique : nombre de gènes identifiés, effets des mutations, expression
des gènes en fonction du milieu, avec ou sans Pi.
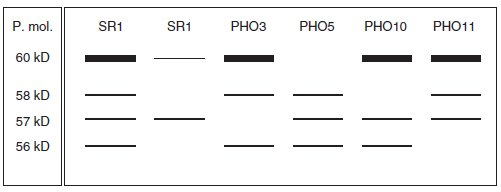
Figure 10.4
– Profils électrophorétiques des activités phosphatases acides en fonction
des différentes souches et de leur milieu de culture.
– À gauche, poids moléculaires correspondant aux différentes bandes, en
dessous dosage de l’activité spécifique.
– Souche (de gauche à droite) : SR1 SR1 PHO3 PHO5 PHO10 PHO11
– Milieu de culture : sans Pi avec Pi sans Pi sans Pi sans Pi sans Pi
– Activité Spécifique : 30 3 25 6 24 23