a. Les deux derniers milieux contiennent de l’arginine et sont destinés à s’affranchir de l’auxotrophie
pour cet acide aminé qui ne concerne que la souche 7.
b. Le but de ce croisement est double :
– le phénotype du diploïde 2n permet de statuer sur la dominance ou la récessivité du phénotype
mutant; ici on peut conclure que tous les phénotypes mutants sont récessifs vis-à-vis
du phénotype sauvage sauf celui du mutant 5.
– l’analyse des fréquences des spores haploïdes issues de la méiose du diploïde d’un tel croisement
permet de réaliser un test de ségrégation 2/2 et de statuer sur le fait que le mutant
croisé par sauvage est un mutant simple ou non.
Si c’est un mutant simple, ne différant de SSR que pour un gène, le diploïde est hétérozygote
pour ce gène et l’analyse des spores doit permettre d’observer le résultat de la ségrégation 2-2
de ce couple d’allèles et de leurs phénotypes associés.
Souches de génotypes (a) × (a+)
Diploïde de génotype (a//a+)
Spores issues de la méiose (a) (a+)
Fréquences des spores 50 % 50 %
Phénotypes des spores [muté] [sauvage]
Si c’est un mutant double, différant de SSR pour deux gènes, le croisement s’écrit alors :
Souches de génotypes (a, b) × (a+, b+)
Diploïde de génotype (a//a+ ----- b//b+)
où les pointillés indiquent notre ignorance quant à une éventuelle liaison génétique entre les
locus des deux gènes
Spores issues de la méiose (a, b) (a+, b+) (a, b+) (a+, b)
Fréquences des spores (1-r)/2 (1-r)/2 r/2 r/2
Phénotypes des spores [muté] [sauvage] [non sauvage] [non sauvage]
Si les deux gènes sont indépendants, on attend 25 % de spores sauvages et si les deux gènes
sont liés, on attend une fréquence comprise entre 25 % et 50 % (fréquence obtenue en cas de
liaison absolue, sans crossing-over, simulant une ségrégation 2/2).
Pour toutes les souches, sauf les souches 3 et 7, on observe que sur le total des spores poussant
sur Mo + iso + val + leu, environ 50 % poussent sur la réplique sur Mo; on peut donc en
conclure qu’il y a ségrégation 2/2 pour le phénotype mutant et que toutes les souches sauf
3 et 7 sont des mutants simples.
Pour la souche 3, on observe 23 spores sauvages, soit environ 25 %, ce qui permet de
conclure qu’il s’agit d’un double mutant avec deux gènes génétiquement indépendants.
Pour la souche 7, on observe 57 spores [arg+] sur un total de 113 (ségrégation 2/2 pour le seul
phénotype arg), ce qui prouve qu’un seul des gènes de la chaîne de biosynthèse de l’arginine
est touché; par ailleurs, sachant que la souche 7 est mutée dans le gène TD, on vérifie par la
ségrégation 2/2 pour le phénotype d’auxotrophie pour l’isoleucine qu’elle n’est mutée que dans ce gène TD (58 spores iso– sur 113).
La souche 7 est donc un double mutant et comme
on obtient 43 spores sauvages [arg+, iso+] sur un total de 113, soit 38 %, cela prouve que les
gènes ARG (gène a) et TD (gène b) sont liés.
On observe 57 – 43 = 14 spores recombinées (a–, b+) de phénotype [arg+, iso–] et 58 – 43 = 15
spores recombinées (a–, b+) de phénotype [arg–, iso+], soit un total de 29 spores recombinées
sur 113 spores testées : le taux de recombinaison est égal à 25,6 % et la distance entre les gènes
ARG et TD est estimée à 25,6 u.r.
c. Test de χ2
Les effectifs observés sont de 23 spores [+], poussant sur Mo et de 52 spores [-] ne pouvant
y pousser.
Sous l’hypothèse nulle d’une ségrégation 2-2 résultant du fait que les souches
parentales ne différeraient que pour un seul gène, les effectifs attendus seraient égaux entre
eux et égaux à 37,5, ce qui conduit à une valeur observée du χ2 égale à 11,2.
Cette valeur observée du χ2 est hautement significative, elle dépasse largement la valeur
seuil, au risque de 5 %, de 3,84 pour un χ2 à 1 degré de liberté.
On peut donc exclure l’hypothèse
de ségrégation 2-2 avec un risque d’erreur, dans la décision, très largement inférieur à
5 % et admettre que la souche 3 est mutée pour plus d’un gène.
Question 4.
a. Le but de ces croisements est de réaliser un test de complémentation fonctionnelle et
d’allélisme, quand il s’agit de croisements entre mutants récessifs.
Si l’un des mutants est dominant, le diploïde est toujours de phénotype mutant et ne peut être
interprété en terme de test de complémentation ou d’allélisme, mais dans tous les cas la
méiose du diploïde permet de tester la liaison entre les mutations.
Le mutant 5 est dominant et exclu de l’analyse fonctionnelle.
L’analyse du tableau pour les autres
mutants permet de définir 6 groupes de complémentation : (2), (3,6), (3,4), (7, 10), (8, 11)
et (9).
Les mutants 4 et 6 complémentent et ne sont donc pas mutés dans le même gène (ce sont des
mutants simples, voir plus haut), on retrouve le fait que 3 est muté dans deux gènes, ce qui
confirme bien les conclusions précédentes.
Question 5.
– Les mutants 2 et 4 sont des mutants simples touchés dans deux gènes différents (puisqu’ils
complémentent), les effectifs observés, 25 % de spores sauvages permettent de conclure
que ces deux gènes sont génétiquement indépendants.
– Les mutants 2 et 9 sont des mutants simples touchés dans deux gènes différents (puisqu’ils
complémentent), les effectifs observés, 1 % de spores sauvages permettent de conclure que
ces deux gènes sont génétiquement liés, avec une distance égale à 2 u.r. (2 % de spores
recombinées, dont la moitié sont sauvages).
Problème 10.3
Dans tout le problème on considère qu’on dispose, pour les croisements,
des souches adéquates, porteuses du bon signe sexuel ainsi que des marqueurs
de sélection des diploïdes.
On a obtenu, de manière indépendante, chez la levure Saccharomyces cerevisiae,
11 mutants incapables d’utiliser le galactose comme source de carbone
et d’énergie, phénotype noté [gal–].
Ces mutants, nommés m1, m2, …, m11, sont par ailleurs auxotrophes pour l’histidine (incapables d’assurer la synthèse
de cet acide aminé), phénotype noté [his–].
On croise chacun des mutants avec une souche, nommée S, de phénotype
[gal+; trp–], capable d’utiliser le galactose mais auxotrophe pour le tryptophane.
Question 1.
a. Quel est le milieu de culture de la boîte sur laquelle on cultive les souches
mutantes et la souche S ?
b. Quel est le milieu de culture de la boîte sur laquelle on réalise un croisement
mi × S ?
Justifiez votre réponse en précisant le rôle des mutations
d’auxotrophie.
Question 2.
a. De quelle manière est testé le phénotype des diploïdes obtenus précédemment
vis-à-vis de leur capacité à utiliser le galactose ?
b. Tous les diploïdes obtenus sont [gal+]; concluez (deux lignes).
Question 3.
a. Le diploïde issu du croisement m1 × S est mis à sporuler et on teste
1 000 spores haploïdes issues de la méiose, parmi lesquelles 525 sont [gal+]
et 475 sont [gal–]; quelle conclusion en tirez-vous ?
On demande une
analyse s’appuyant sur la formulation de génotypes.
b. Le même type de résultat est obtenu quand on étudie les spores haploïdes
formées à la méiose par les diploïdes issus des croisements entre S et chaque
mutant mi, sauf m10; concluez (une ligne).
Question 4.
On étudie un échantillon important de spores haploïdes issues du diploïde
m10 × S, assez important pour considérer comme significativement différentes
les fréquences de spores [gal–] et [gal+], respectivement égales à
54 % et 46 %.
Quelle est l’hypothèse génétique la plus simple pour expliquer un tel résultat ?
On demande une analyse s’appuyant sur la formulation de génotypes.
Question 5.
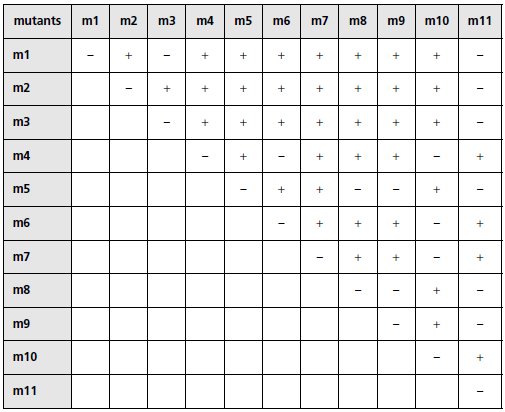
On croise les mutants entre eux et on teste le phénotype des diploïdes pour
le galactose (tableau ci-dessous où « + » désigne le phénotype [gal+] et « – »
désigne le phénotype [gal–]).
a. Interprétez, en justifiant votre réponse, les observations rapportées par
ce tableau par le regroupement des mutants dans des groupes adéquats.
b. Montrez en quoi le résultat concernant le mutant m10 sont cohérents
avec les conclusions de la question 4.
c. Quelle hypothèse vous suggère le résultat concernant le mutant m11 si
vous considérez, en même temps, le résultat observé pour ce mutant dans
la question 3-b ?
Question 6.
On dispose d’un autre mutant de levure, noté B, de phénotype [ade–; met–],
auxotrophe pour l’adénine et la méthionine.
Le croisement de B avec une souche sauvage [ade+; met+] donne des
diploïdes à partir desquels on obtient, après méiose, 1 000 spores haploïdes,
dont la répartition est donnée ci-dessous.
Faites l’analyse génétique complète
de ces résultats (nombre de gènes, indépendance ou liaison, etc.).
On demande
une analyse s’appuyant sur la formulation de génotypes.
[ade+; met+] = 310 [ade–; met–] = 290
[ade–; met+] = 205 [ade+; met–] = 195
Question 7.
Le croisement de B avec le mutant m11 donne des diploïdes à partir desquels
on obtient, après méiose, 1 000 spores haploïdes se répartissant ainsi :
[ade+; met+] = 360 [ade–; met–] = 340
[ade–; met+] = 140 [ade+; met–] = 160
Montrez que la différence entre le résultat observé dans le croisement B × m11
ci dessus et celui observé à la question 6 vous permet de confirmer l’une
des hypothèses faites à la question 5-c.
Il est inutile de reprendre ici toute
l’analyse génétique faite à la question 6, mais seulement un des points
de cette analyse.
Question 8.
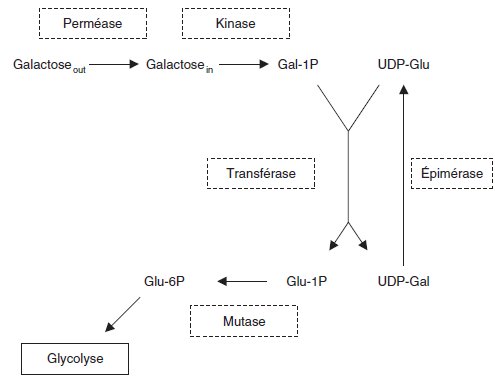
Des études biochimiques ont montré que la métabolisation du galactose
dépendait de cinq gènes de structure dont les fonctions permettent l’entrée
du galactose puis sa transformation en Glucose6-Phosphate pour intégrer le
métabolisme énergétique par la glycolyse par la voie D’Emden-Meyerhoff,
selon le schéma ci-dessous :
L’analyse biochimique, par dosage des activités ou tests de perméation chez
les différents des mutants a permis de dégager les résultats rapportés par le
tableau suivant où « + » désigne la présence de l’activité et « – » son absence.
En reprenant vos résultats des questions précédentes, on vous demande de :
a. Montrer la cohérence pour les mutants m1, m2, m3, m5, m7 et m11,
entre les observations génétiques (questions 2 et/ou 5) et les observations
fonctionnelles rapportées par le tableau ci-dessus, sachant qu’on a montré, par ailleurs, que les gènes de structure de la kinase, de l’épimérase et de la
transférase étaient contigus.