Le type sauvage d’une espèce du genre Sordaria produit des spores de
couleur noire [n], dans un asque à spores ordonnées mais non orientées
(ni haut, ni bas). L’asque contient huit spores (octade) à la suite d’une
mitose additionnelle survenant après la méiose II.
1. On dispose de deux mutants l’un à spores jaunes [j], l’autre à spores
roses [r].
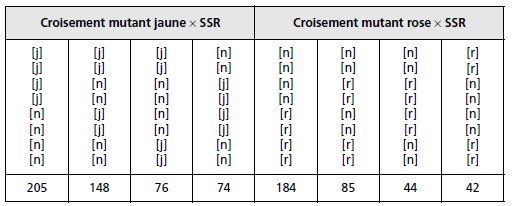
Chacun des deux mutants est croisé avec le type sauvage, puis on observe
les asques issues de la méiose (tabl. 4.6). Interprétez ces résultats.
TABLEAU 4.6.
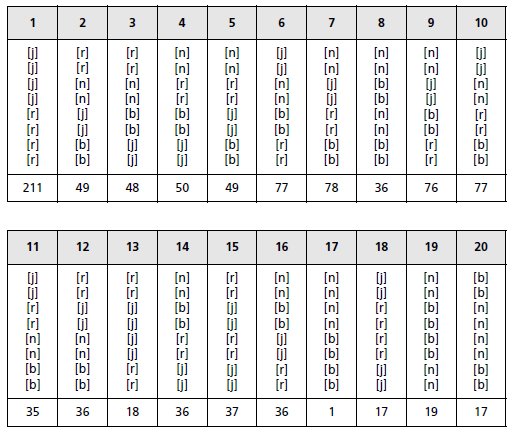
2. On croise les deux souches mutantes, et on observe 20 types différents
de tétrades (tabl. 4.7). [b] correspond à un phénotype de spore blanche.
Interprétez ces résultats de manière exhaustive, fonctionnelle et cartographique.
TABLEAU 4.7.
3. Montrez en quoi le résultat quantitatif concernant l’asque de type 17
prouve que la survenue d’un crossing-over, sur un bras chromosomique,
bloque la survenue d’un deuxième crossing-over, sur ce même bras (interférence
positive); il n’y aurait donc au plus que deux crossing-over par
chromosome, un de chaque côté du centromère.
Pour cela il est utile de schématiser les deux paires de chromatides en
prophase I et d’en déduire les conséquences génétiques selon qu’il y a 0, 1
ou 2 crossing-over afin de définir, dans chacun des 20 types d’asques
observés, pour lequel des gènes en jeu, il y a eu pré ou postréduction,
notamment pour l’asque de type 17.
– Maîtriser les mécanismes de la méiose, de la ségrégation 2/2 à l’analyse de
l’indépendance ou de la liaison génétique, à travers de tétrades.
– Distances gène-centromère et distances entre gènes.
– Approche fonctionnelle de la diversité phénotypique associée au mode de
transmission.
– Analyse de la pré et de la postréduction pour des gènes liés, interférence entre
crossing-over.
Solution
1. Analyse de ségrégation.
Dans chacun des deux croisements, les asques renferment 4 spores
de type sauvage et 4 spores de type mutant, ce qui illustre le résultat attendu d’une ségrégation
2/2 d’un couple d’allèles à la méiose.
On peut considérer que le mutant à spores jaunes
diffère du type SSR pour un seul gène, dont les allèles seront notés J chez la SSR et j chez le
mutant (remarque 1, ex. 4.1) et que le mutant à spores roses diffère du type SSR pour un seul
gène, dont les allèles seront notés R chez la SSR et r chez le mutant.
Cartographie par rapport au centromère.
Les octades étant ordonnées, il est possible de
distinguer les méioses préréduites, conduisant à deux demi-asques homogènes, des méioses
postréduites conduisant à deux demi-asques hétérogènes.
Dans les deux cas, la fréquence de postréduction est significativement inférieure à sa limite
de 0,666, atteinte dès qu’il y a ségrégation indépendante entre le locus du gène et son centromère.
On peut donc estimer la distance au centromère de chacune des mutations j ou r par la
demi-fréquence de postréduction multipliée par 100 (remarque 2, ex. 4.1), soit :
– distance de j à son centromère : dj = 29,5 urp (unités de recombinaison par postréduction);
– distance de r à son centromère : dr = 24 urp.
Remarque.
Il est difficile, à ce stade d’émettre quelque avis sur le plan fonctionnel.
Il
est peu aisé d’envisager, pour un phénotype spécifique de la spore haploïde, un test de
complémentation fonctionnelle !
On pourrait conclure que les mutations j et r ne sont pas alléliques si on montrait
qu’elles touchent des gènes physiquement indépendants, d’où l’importance du croisement
entre souches mutantes pour en étudier les produits de la recombinaison génétique
à la méiose.
2. Le croisement entre les deux souches jaunes et roses peut s’écrire formellement, ( j; R)
× (J; r), ce qui donne un diploïde (j//J; R//r) si les gènes sont physiquement indépendants et
un diploïde ( jR//Jr) s’ils sont physiquement liés. Plusieurs résultats doivent être interprétés.
• Apparition de spores noires et de spores blanches. Bien évidemment les spores noires sont
de phénotype et de génotype sauvage (J; R) et sont le résultat d’une recombinaison génétique
entre les deux apports parentaux.
On observe qu’à la présence d’une spore noire recombinée, est toujours associée la présence
d’une autre spore de phénotype blanc correspondant obligatoirement au génotype recombiné
réciproque (j; r).
On peut donc faire l’hypothèse fonctionnelle que les mutations j et r touchent deux gènes soit
dans une même voie de biosynthèse du pigment noir, les précurseurs accumulés (ou leurs
dérivés) chez les mutants étant jaunes ou roses, soit dans deux voies parallèles.
Rien ne peut
être précisé à ce stade, sauf le fait que le double mutant bloque la formation de tout pigment
d’où la couleur blanche.
Remarque. On a clairement démontré que le double mutant confère le phénotype
blanc, mais pas vraiment qu’il est muté dans deux gènes différents.
Cependant il est
difficile de donner une interprétation fonctionnelle simple de l’apparition de phénotypes
différents de pigmentation selon qu’un seul gène serait muté en un site ou un
autre ou aux deux, d’autant que sous cette hypothèse les deux sites mutés seraient trop
distants pour être sur un même gène (29,5 – 24 = 5,5 urp au minimum si les gènes
sont liés et du même côté du centromère, voir question précédente); mais on connaît
l’existence de points chauds de recombinaison qui, en augmentant considérablement
la probabilité locale de crossing-over, peuvent faire paraître génétiquement éloignés
deux points qui sont physiquement très proches.
• Analyse de la recombinaison génétique entre les deux gènes, puis de la pré ou de la postréduction
pour chacun des deux gènes.
L’interprétation des 20 types d’asques permet de
savoir si ce sont des ditypes parentaux (DP) avec des spores parentales jaunes ou roses, ou
des ditypes recombinés (DR), avec des spores recombinées noires ou blanches, ou des tétratypes
(T), avec les quatre types de spores (tabl. 4.8).
TABLEAU 4.8 DESCRIPTION DES 20 TYPES D’ASQUES ET EFFECTIFS OBSERVÉS.
Rappel : il y a 36 types d’asques possibles pour des tétrades ordonnées avec orientation
et 20 types quand il n’y a pas d’orientation.
Les trois types d’asques, DP, DR et T ont des fréquences respectivement égales à 0,282;
0,073 et 0,645.
La fréquence des DP étant significativement supérieure à celle des DR, on
peut conclure à la liaison génétique et physique des deux locus et estimer une distance génétique
corrigée.
La distance corrigée entre les mutations j et r est égale à : djr = [f(T)/2 + 3f(DR)] × 100 = 54 ur
qui se trouve remarquablement cohérente avec la somme des distances de chacune des mutations
au centromère, celui-ci se trouvant central, ce qui montre définitivement que les deux
mutations j et r touchent deux gènes différents, puisque situés de part et d’autre d’un
centromère.
L’additivité presque parfaite des distances estimées trouve son explication dans la question
suivante.
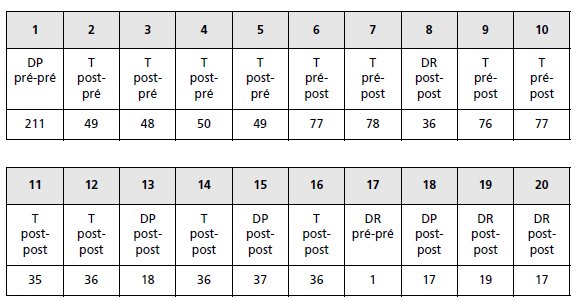
3. La reconstitution des génotypes pour chaque type de spore permet de savoir pour chacun
des deux gènes s’il est pré ou postréduit (asques homogènes ou hétérogènes pour les deux
allèles de chacun des deux gènes).
D’où le bilan présenté dans le tableau 4.8 où le premier
terme (pré ou post) se réfère au couple R//r et le deuxième au couple J//j.
Pour obtenir des DR, il est nécessaire de réaliser deux crossing-over entre les quatre chromatides
(par exemple, 1-4 pour un crossing-over et 2-3 pour l’autre, ou bien 1-3 et 2-4, voir
rappels de cours).
Comme les deux gènes sont de part et d’autre du centromère, on peut obtenir des DR, soit
avec deux crossing-over d’un même côté du centromère et aucun de l’autre côté, soit avec un
crossing-over de chaque côté du centromère; dans le premier cas on obtiendra des DR prépré
(préréduits pour chacun des deux gènes), dans le second cas on obtiendra des DR postpost.
On observe 1 DR pré-pré pour 72 DR post-post; la faible fréquence des DR pré-pré indique
que la survenue d’un deuxième crossing-over sur le même bras chromosomique qu’un
premier est un événement très rare (dans cette espèce).
Les DP pré-pré ou les T post-pré, obtenus par doubles crossing-over sur un même bras, sont
sans doute aussi rares mais ne peuvent être distingués des DP obtenus sans crossing-over qui
sont eux aussi pré-pré, ou des T obtenus avec un seul crossing-over qui sont eux aussi postpré.
Remarque. Une fois encore, ce sont les DR qui permettent d’apporter une information
décisive sur les événements survenus à la méiose.