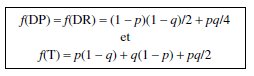De la même manière que la fréquence de gamètes recombinés entre deux gènes
physiquement liés dépend de leur distance, la fréquence des asques post-réduits
dépendra de la distance entre le locus du gène et son centromère.
Aussi, de la même manière qu’on peut estimer la distance entre deux gènes (génétiquement
liés) à travers leur fréquence r de recombinaison, de la même manière, il
sera possible d’estimer la distance d’un gène à son centromère à travers sa fréquence p
de post-réduction, à condition que la valeur de p soit inférieure à une certaine valeur,
de la même façon que r doit être inférieure à ½ pour qu’on puisse conclure à une
liaison génétique et calculer une distance.
Nous allons montrer que la valeur limite de
p est égale à 2/3, alors que la valeur limite de r est égale à ½.
En effet p, la fréquence de post-réduction n’est pas égale à la probabilité qu’un
CO survienne entre le locus du gène et son centromère, car il peut, quand le gène est
assez éloigné, survenir deux CO qui, selon les chromatides impliquées, peuvent
conduire soit à une pré-réduction, soit à une post-réduction.
Ainsi p est une fréquence
(ou une probabilité) qui tient compte d’une multitude d’événements possibles et non
du seul événement d’un CO entre le gène et son centromère.
• Si la distance du locus du gène au centromère est nulle, aucun crossing-over ne
survient dans aucune méiose, et la valeur de p est nulle.
En inversant la proposition, on peut conclure d’une absence de postréduction (sur
un grand nombre de méioses) que la distance du gène à son centromère est nulle, ou,
en tout cas, suffisamment faible pour qu’aucun crossing-over ne soit survenu sur le
nombre de méioses étudiées.
• Si la distance du locus du gène à son centromère est telle qu’un crossing-over au
plus peut survenir dans quelques méioses, on peut mesurer la distance à partir du
taux p de postréduction, par analogie au calcul de la distance génétique en unités de
recombinaison.
La distance génétique en unités de recombinaison est égale à la
fréquence de gamètes recombinés multipliée par cent.
La fréquence des
gamètes recombinés étant une visualisation ou une mesure plus ou moins exacte des
chromatides remaniés par un crossing-over entre les locus des deux gènes.
Par analogie, la distance D entre le locus d’un gène et son centromère est estimée
par la fréquence (multipliée par cent) des chromatides « remaniées » entre le centromère
et le locus du gène.
Celles-ci ne concernent que la moitié des spores d’un asque postréduit, aussi la fréquence des chromatides remaniées par un crossing-over entre
le gène et son centromère est égale à p/2, d’où :
D = p/2 × 100, en unités de recombinaison par postréduction (urp).
Remarque. On verra plus loin, qu’en toute rigueur il ne faut pas confondre les
unités de recombinaison par crossing-over (notées ur) et celles par postréduction
(urp).
• Si la distance du locus du gène à son centromère est telle qu’au moins un, voire
plusieurs crossing-over peuvent survenir dans une méiose, la postréduction devient
très fréquente et tend vers une limite égale à 2/3, ce qui se démontre aisément :
– quand un gène est très éloigné de son centromère, le nombre de crossing-over est
tel que les quatre exemplaires du gène ségrègent indépendamment les uns des
autres;
– le premier des deux allèles A peut ségréger à la méiose I avec le deuxième allèle
A, événement de probabilité égale à 1/3 (sur les trois allèles restants, seul l’un
d’entre eux est A), ce qui conduit à une préréduction;
– le premier des deux allèles A peut ségréger à la méiose I avec l’un des deux allèles a,
événement de probabilité égale à 2/3 (sur les trois allèles restants, deux d’entre
eux sont a), ce qui conduit à une postréduction.
Cette valeur limite p = 2/3 de la postréduction définit la limite au-delà de laquelle
l’estimation d’une distance d’un gène à son centromère n’est plus possible, puisqu’alors
deux gènes très distants du centromère présenteront tous deux des taux de postréduction
égaux à 2/3, bien que leurs distances au centromère puissent être très différentes
mais qu’elles sont toutes deux suffisamment grandes pour entraîner une ségrégation
indépendante des allèles de chacun des gènes par rapport au centromère.
On ne peut donc estimer la distance d’un gène à son centromère que s’il n’y a pas
de ségrégation indépendante entre le centromère et les allèles de ce gène; donc si la
fréquence p de postréduction est strictement inférieure à 2/3.
Remarque 1. La valeur limite de l’estimation de la distance au centromère
est donc égale à 33 urp (2/3 × 1/2 × 100) ce qui n’est pas la limite de la
distance de recombinaison entre deux gènes par crossing-over, égale à 50 ur
(1/2 × 100).
Cette différence vient de ce que le centromère ne se comporte pas
du tout comme un gène. En effet, il n’y a jamais de disjonction à la méiose I
pour un centromère joignant des chromatides soeurs, alors qu’il peut y avoir,
en raison des crossing-over, une disjonction des allèles des gènes portés par
ces mêmes chromatides soeurs !
Remarque 2. Tout comme la distance génétique de recombinaison, la distance
cartographique au centromère est bien estimée quand elle est suffisamment
petite pour qu’un crossing-over au plus survienne entre le gène et son centromère,
puis devient sous-estimée dès que deux crossing-over peuvent survenir.
En effet, dans ce cas, les deux crossing-over peuvent affecter les mêmes chromatides,
ou les quatre chromatides, et redonner de la préréduction, la mesure
de la postréduction ne permettra pas de comptabiliser ces asques préréduites
dont certaines spores sont pourtant porteuses de chromatides remaniées, et
même doublement remaniées.
Remarque 3. Chez des ascomycètes comme Saccharomyces cerevisiae
(levure de boulangerie) ou Aspergillus nidulans (moisissure verte du pain), les
tétrades sont inordonnées, et il n’est pas possible de ce fait, d’analyser la pré et la postréduction, ou de cartographier les gènes par rapport à leur centromère,
du moins directement, car cela est possible indirectement avec des
marqueurs centromériques (voir exercices).
L’étude
de l'indépendance et de la liaison génétique par l'analyse de
tétrades :
L’analyse de tétrades, pour une méiose affectant deux gènes représentés par deux
couples d’allèles, permet de définir toutes les configurations possibles de méioses
inaccessibles dans l’étude de gamètes en vrac et de préciser certains aspects de la
méiose souvent méconnus ou trop simplifiés.
A - Analyse de tétrades pour deux gènes physiquement
indépendants :
a) Trente-six configurations possibles de la méiose
:
Le croisement d’une souche haploïde {A; B} par une souche haploïde {a; b} donne
une cellule diploïde {A//a; B//b}.
Pour chacun des deux gènes on a (voir plus haut) six
configurations possibles, deux préréduites et quatre postréduites, et sous l’hypothèse
qu’une paire d’homologues ségrège indépendamment d’une autre paire d’homologues,
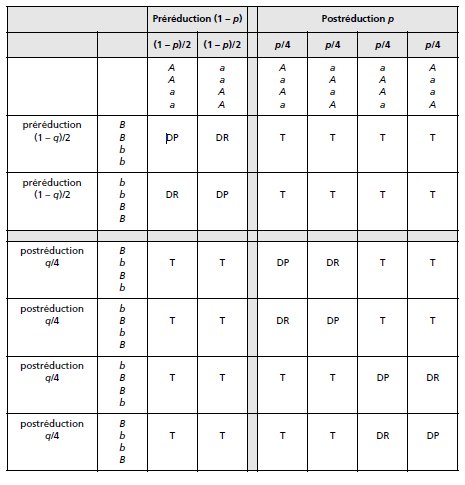
on peut prévoir 6 × 6 = 36 configurations possibles non superposables (tabl. 4.1).
Dans un organisme où survient une mitose supplémentaire le stock des spores est
doublé, sans que cela change la configuration; on s’abstiendra donc de cette mitose.
Les taux de postréduction pour les couples d’allèles A/a et B/b sont respectivement
figurés par p et q.
L’analyse de tétrades totalement ordonnées (avec haut et bas), pour deux gènes
physiquement indépendants, permet effectivement l’observation de ces 36 types
d’asques non superposables, avec des fréquences égales aux valeurs prévues sous
l’hypothèse d’une ségrégation indépendante des paires d’homologues, soit le produit
des fréquences marginales fonction de p et q.
La cohérence entre prévisions et observations constitue bien une validation expérimentale
directe du fait, qu’à la méiose, les paires d’homologues différents ségrègent
indépendamment l’une de l’autre.
Remarque. Si on dispose d’asques ordonnés sans haut ni bas, les asques sont
identiques deux à deux, sauf le dernier carré en bas à droite du tableau, ce qui
réduit à vingt le nombre de configurations différentes de la méiose.
b) Définition des ditypes parentaux ou recombinés et des tétratypes
Les 36 types d’asques peuvent être, si on ne tient plus compte de l’ordre des spores
dans l’asque, regroupés en trois types d’asques :
– les ditypes parentaux (DP) avec quatre spores, deux à deux identiques à chacun
des deux types parentaux, ici {A; B} et {a; b};
TABLEAU 4.1 CONFIGURATIONS POSSIBLES DE LA MÉIOSE.
– les ditypes recombinés (DR) avec quatre spores, deux à deux identiques à chacun
des deux types recombinés possibles, ici {A; b} et {a; B};
– les tétratypes (T) avec quatre spores, toutes de génotypes différents, deux de type
parental, ici {A; B} et {a; b}, et deux des deux types recombinés possibles, ici
{A; b} et {a; B}.
Remarque 1.
La méiose donne systématiquement un tétratype quand un seul
des deux gènes est postréduit.
Le fait qu’une méiose puisse donner quatre
gamètes différents est souvent occulté dans les schémas simplistes de la
méiose (fig. 3.1) qui, n’envisageant pas (par simplicité légitime) la possibilité
de crossing-over entre un gène et son centromère, concluent faussement que
la méiose de deux gènes physiquement indépendants donne soit quatre gamètes parentaux soit quatre gamètes recombinés (DP ou DR du premier
carré en haut à gauche, correspondant à la double préréduction).
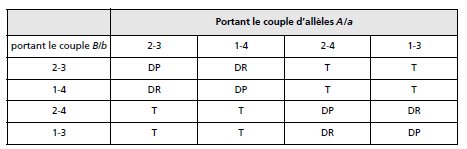
Remarque 2. Quand les deux gènes sont postréduits, on aura des DP, des DR
ou des T, en fonction des paires de chromatides impliquées par chacun des
crossing-over sur chacune des paires d’homologues (tabl. 4.2, on peut obtenir
les résultats rapportés en détaillant les dispositions métaphasiques sur des
schémas à faire à titre d’exercice) :
➤ on obtient des DP si les deux crossing-over impliquent, sur chaque paire
d’homologues, des paires de chromatides non-soeurs situées dans deux
mêmes plans différents (ex. 1-4 puis 1-4);
➤ on obtient des DR si les deux crossing-over impliquent, sur chaque paire
d’homologue, des paires de chromatides non-soeurs situées dans quatre
plans différents (ex. 1-4 puis 2-3);
➤ on obtient des T si les deux crossing-over impliquent, sur chaque paire
d’homologue, des paires de chromatides non-soeurs situées dans trois plans
différents (ex. 1-4 puis 1-3).
TABLEAU 4.2 TYPES D’ASQUES EN FONCTION DE LA POSITION DES CHROMATIDES IMPLIQUÉES
PAR UN CROSSING-OVER SUR LA PAIRE D’HOMOLOGUES.
c) Fréquences des types d’asques et de gamètes
Il est aisé, à partir du tableau des 36 configurations, de montrer que :
Compte tenu de la composition de chaque type d’asques, en gamètes parentaux
(GP) et/ou recombinés (GR), il est facile de montrer que :
On retrouve évidemment, avec l’analyse de tétrades, le résultat connu que deux
gènes physiquement indépendants sont génétiquement indépendants (équifréquence
des gamètes parentaux et recombinés).
d) Valeurs limites des fréquences des trois types d’asques
:
Avec ses trois classes d’asques, l’analyse de tétrades fournit une information plus
riche que l’analyse génétique habituelle avec ses deux classes de gamètes.
De cette information plus riche, on peut tirer souvent des conclusions cartographiques,
comme l’indépendance physique de deux gènes, inaccessibles par l’analyse
des classes gamétiques (voir plus loin).
Le traitement de cette information suppose
de connaître le domaine de variation des fréquences des trois types d’asques.
• Si les deux gènes sont très proches de leurs centromères respectifs, ils ne seront
jamais postréduits et le tableau se réduit au carré des quatre double préréductions, en
haut à gauche du tableau des 36 configurations.
On a alors :
ce qu’on obtient aussi en prenant zéro pour valeurs de p et de q dans les équations
donnant les fréquences des trois types de tétrades.
• Si un seul des deux gènes est génétiquement indépendants de son centromère, de
sorte qu’il présente un taux de postréduction égale à 2/3, les fréquences des trois
types de tétrades deviennent indépendantes du taux de postréduction de l’autre gène,
et sont égales à :
On peut ainsi définir les domaines de variation des fréquences des trois types de
tétrades de la manière suivante :
Remarque. L’analyse des tétrades inordonnées, chez la levure Saccharomyces
cerevisiae ne permet pas, dans l’étude d’un gène A d’identifier les asques postréduits
et d’en tirer l’estimation de la distance du gène au centromère.
Mais si on dispose d’un deuxième gène B connu pour être très proche de son
centromère, on peut faire une analyse de tétrades, pour les deux gènes A et B,
qui conduira à l’estimation de la fréquence des tétratypes.
Comme il ne peut y avoir de crossing-over entre le gène B et son centromère,
on peut conclure que tous les tétratypes dérivent d’une postréduction pour le
gène A.
La fréquence des tétratypes pour A et B est donc en même temps la
fréquence de postréduction pour A. On en tire donc la distance de A à son
centromère; le gène B jouant le rôle de marqueur centromérique coségrégeant
avec son centromère (voir exercices plus loin).