Le batracien Rana pipiens, fréquemment utilisé comme matériel par les
embryologistes, présente aussi un intérêt pour les généticiens. Son cycle de
reproduction est diplobiontique et, comme chez la quasi-totalité des vertébrés,
il est normalement impossible d’obtenir, de manière groupée, les
quatre produits d’une même méiose.
À la ponte de l’oeuf, la première division méiotique a déjà eu lieu; l’un des
deux noyaux a été expulsé dans le premier globule polaire, tandis que
l’autre noyau est entré en méiose II et reste bloqué en métaphase.
Lors de la fertilisation de l’ovocyte, la pénétration du noyau provenant du
spermatozoïde déclenche la sortie de métaphase.
L’anaphase puis la télophase
aboutissent à la formation de deux noyaux haploïdes dont un est
expulsé en un deuxième globule polaire, et l’autre va fusionner avec le
noyau d’origine mâle pour former le zygote, à l’origine d’un nouvel
individu.
Du fait que la fécondation, puis le développement, sont externes, il est
facile de manipuler l’oeuf et l’embryon de batracien.
Les embryologistes
ont découvert un moyen d’activer artificiellement l’ovocyte, sans fertilisation,
ce qui conduit, après l’expulsion du deuxième globule polaire, au
développement d’un embryon haploïde n’arrivant pas au stade adulte.
Puis
ils ont découvert un moyen, après activation artificielle de l’ovocyte,
d’inhiber l’expulsion du deuxième globule polaire, ce qui conduit alors à la
fusion des deux noyaux issus de la méiose II et au développement,
jusqu’au stade adulte, d’individus dénommés « diploïdes gynogéniques »,
puisque tous leurs chromosomes sont d’origine maternelle.
On dispose de deux souches pures de Rana pipiens, l’une de couleur verte,
l’autre de couleur jaune, dont des études génétiques ont montré qu’elles ne
différaient que pour un seul gène (dont les allèles seront notés A et a).
Leur
croisement donne des individus F1 de couleur jaune.
À partir de femelles F1, on obtient des femelles F2 gynogéniques, 58 %
d’entre elles sont jaunes, 42 % sont vertes.
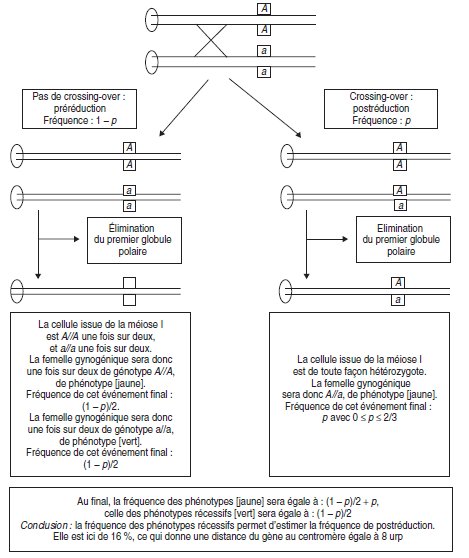
1. Vous nommerez le phénomène génétique visualisé par cette étude, et
vous en ferez un schéma explicite.
2. Vous en déduirez la conséquence cartographique.
– Illustrer l’existence de conditions particulières permettant l’analyse de la pré et
de la postréduction chez un vertébré.
– Distance gène-centromère.
Solution. Il s’agit d’un cas exceptionnel qui permet de visualiser, chez un organisme diplobiontique,
les conséquences génétiques de la pré ou de la postréduction survenant à la méiose
(fig. 4.18).
En conséquence il est possible d’estimer le taux de postréduction (ici 16 %) et d’en déduire
la distance du locus du gène étudié au centromère, ici 8 urp.
Figure 4.18.
Exercice 4.6
:
La levure Saccharomyces cerevisiae est un ascomycète haplodiplobiontique
se présentant sous forme de cellules isolées dans un milieu de culture
liquide et formant des colonies sur un milieu solide.
Les cellules entrant en
méiose produisent quatre spores haploïdes réunies dans un asque inordonné.
On croise entre elles une souche de génotype (Mat a, met), auxotrophe
pour la méthionine, et une souche de génotype (Mat α, trp, ade), auxotrophe
pour les tryptophane et l’adénine.
On rappelle que le croisement
entre deux souches est conditionné par le fait qu’elles soient de signes
sexuels opposés, notés Mat a et Mat α.
Après méiose, on isole 120 tétrades, puis on les décortique sous une loupe
binoculaire pour aligner les spores sur un milieu complet afin d’obtenir des
colonies à partir de chaque spore haploïde.
Ces colonies sont repiquées sur
différents milieux dans le but de tester leurs génotypes, le signe sexuel
étant testé par croisement avec une souche de signe Mat a ou Mat α.
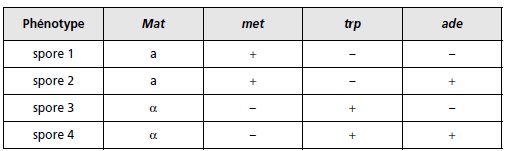
1. Caractérisez le type de tétrade obtenue, ditype parental (DP), ditype
recombiné (DR) ou tétratype (T), pour chaque couple de phénotypes, dans
l’analyse des quatre spores d’une tétrade (tabl. 4.9).
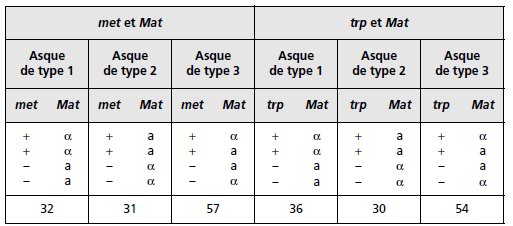
TABLEAU 4.9 RÉSULTATS OBTENUS POUR L’UNE DES TÉTRADES.
Le signe « + » indique que la spore est prototrophe pour le phénotype
considéré, et le signe « – » qu’elle est auxotrophe.
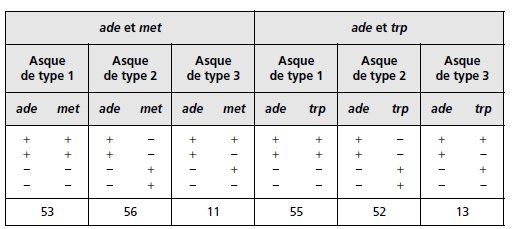
2. On analyse 120 tétrades, et on regroupe les résultats pour chacun des
gènes pris deux à deux (tabl. 4.10). Interprétez ces résultats de manière
exhaustive, en calculant toutes les distances génétiques, y compris celles
par rapport au centromère.
TABLEAU 4.10. Le signe « + » indique que la spore est prototrophe pour le phénotype
considéré, et le signe « – » qu’elle est auxotrophe.
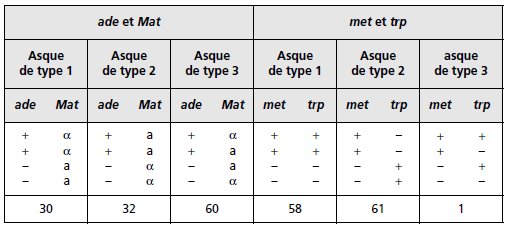
TABLEAU 4.10 (SUITE).
TABLEAU 4.10 (SUITE).
– Analyse de tétrades chez la levure Saccharomyces cerevisiae.
– Distance gène-centromère par utilisation de marqueurs centromériques.
Solution
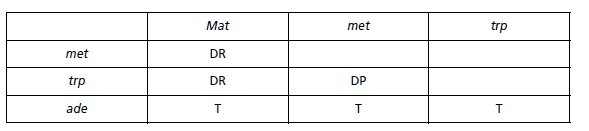
1. On définit l’asque comme DP, DR ou T relativement à deux phénotypes ou deux gènes
gouvernant ces deux phénotypes.
Un asque peut très bien être DP pour deux gènes alors qu’il
sera DR ou T pour deux autres, ce qui est le cas ici, puisqu’il suffit de considérer les gènes
deux à deux pour voir si les spores sont de type parental ou recombiné, ce qui conduit à :
TABLEAU 4.11.
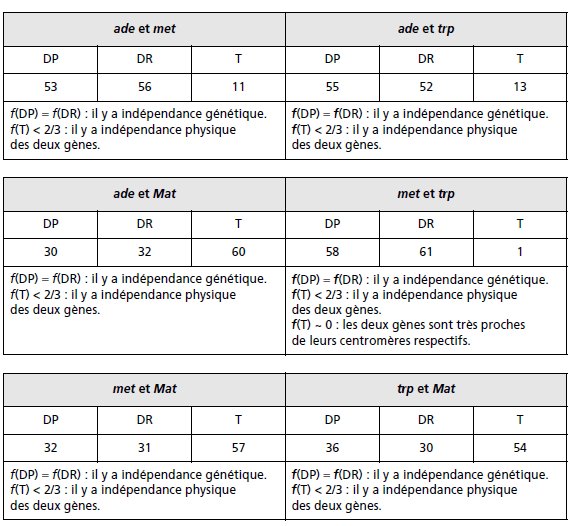
2. Pour chacun des couples de gènes on peut définir et comptabiliser le type d’asque observé,
le type 1 étant les DP, le type 2 étant les DR, et le type 3 étant les T.
On peut alors donner
pour chaque couple de gènes la conclusion cartographique résultant de l’analyse des
fréquences des DP, des DR et des tétratypes (tabl. 4.12), compte tenu de l’algorithme de décision
défini (fig. 4.10).
Comme les deux gènes met et trp sont proches de leurs centromères respectifs, on peut considérer
qu’ils ne sont jamais postréduits et que coségrégeant avec leur centromère, ils en constituent
un « marqueur ».
En conséquence, dans les croisements ade × met ou ade × trp, les
tétratypes résultent exclusivement de la postréduction pour le gène ade, ce qui permet alors,
bien que les tétrades ne soient pas ordonnées, d’estimer, grâce au marqueur centromérique
que sont met ou trp, la distance de ade à son centromère.
Pour le gène ade, on a la fréquence de postréduction égale à celle des tétratypes observés
avec les deux marqueurs centromériques, car il est statistiquement meilleur de prendre la
somme des observations, soit p = 24/240, et la distance au centromère qui est égale à
d = (p/2) × 100 = 5 urp.
Pour le gène Mat, on observe p = 111/240 = 0,46, ce qui donne une distance au centromère
égale à 24 urp.
TABLEAU 4.12 CONCLUSIONS DE L’ANALYSE GÉNÉTIQUE DES GÈNES PRIS DEUX À DEUX.
On remarquera la cohérence des résultats puisque dans le croisement Mat × ade, on attend
que les tétratypes aient une fréquence donnée par l’équation f(T) = p(1 – q) + q(1 – p) + pq/2
(voir page 104), où p (0,1) et q (0,46) sont les fréquences de postréduction pour les deux
gènes étudiés (voir le rappel de cours), soit une valeur f(T) = 0,491, ce qui représente sur
120 tétrades, un effectif théorique de 58,92 tétratypes; on en observe 60 !