Le type sauvage d’une espèce du genre Sordaria produit des spores de
couleur noire, dans un asque à spores ordonnées mais non orientées (ni haut,
ni bas). L’asque contient huit spores (octade) à la suite d’une mitose additionnelle
survenant après la méiose II.
On croise un mutant à spores vertes avec une souche SSR, et on observe, à
l’issue des méioses, quatre types d’asques (tabl. 4.3). Faire l’analyse génétique
complète de ces résultats.
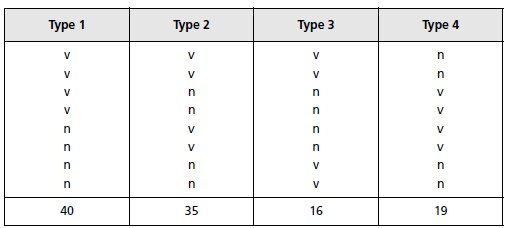
TABLEAU 4.3.
Maîtriser les mécanismes de la méiose et leurs conséquences génétiques par la
reconstruction de tous les scénarios ayant conduit à une situation donnée, prendre
en compte les spécificités de Sordaria.
Solution. Dans ce croisement, les asques renferment 4 spores de type sauvage et 4 spores de
type mutant, ce qui illustre le résultat attendu d’une ségrégation 2/2 d’un couple d’allèles à la
méiose.
On peut considérer que le mutant à spores vertes diffère du type SSR pour un seul
gène, dont les allèles seront notés V chez la SSR et v chez le mutant.
Le diploïde est V//v et
donne, à la méiose, 50 % de spores (V) et 50 % de spores (v).
Remarque 1. Il n’a pas été dit que le phénotype vert n’était gouverné que par un gène,
car on peut imaginer que des mutations dans plusieurs gènes différents puissent donner
le même phénotype vert !
Le mutant étudié est muté dans l’un (s’il y en a plusieurs) des gènes impliqués dans le
phénotype vert.
Une ségrégation 2/2 montre que deux souches croisées entre elles ne diffèrent, pour le
phénotype étudié, que pour un seul gène, mais cela ne signifie nullement que le
phénotype étudié ne dépend que de ce seul gène.
Cartographie par rapport au centromère.
Les octades étant ordonnées, il est possible de
distinguer les méioses préréduites, conduisant à deux demi-asques homogènes, des
méioses postréduites conduisant à deux demi-asques hétérogènes.
On observe 40 asques préréduits et 70 asques postréduits ; il convient de remarquer que les quatre types d’asques
postréduits équifréquents observables pour les octades ordonnées et orientées (avec bas et
haut) se réduisent à trois, en absence d’orientations des asques (ni bas, ni haut).
Mais alors
les trois types d’asques ne sont plus équifréquents, l’un des trois types (où il y a alternance
des couples de spores) est en fait un cas double correspondant à deux scénarios possibles
et équifréquents de la méiose, donnant deux résultats différents s’il y a orientation, et le
même en l’absence d’orientation (voir les figures pages 98-100).
C’est ce qu’on remarque
avec l’octade de type 2 dont l’effectif est environ le double de celui de chacun des deux
autres.
La fréquence de postréduction (70/110 = 0,64) n’est pas significativement différente de sa
limite de 0,666 atteinte dès qu’il y a ségrégation indépendante entre le locus du gène et son
centromère; on peut donc conclure que les allèles du gène étudié ségrègent indépendamment
du centromère et qu’on ne peut, dès lors, estimer la distance entre le locus du gène et le
centromère.
Remarque 2. La cohérence veut qu’on vérifie la liaison au centromère avant
d’estimer une distance.
Il ne serait pas logique d’estimer une distance et de dire qu’il
y a ségrégation indépendante parce qu’elle est égale à sa valeur limite 33,3 urp (demifréquence
de postréduction multipliée par 100).
En effet, pour tous les locus ségrégeant
indépendamment du centromère, on aura 66 % de postréduction, quelle que soit
leur distance au centromère !
Le même genre de faute logique est fait quand, ayant étudié la méiose pour deux
gènes et estimé une fréquence de gamètes recombinés égale à 50 %, on estime une
« distance » de 50 ur pour conclure qu’il y a indépendance génétique.
C’est même ici
plus grave puisque les deux gènes peuvent être physiquement indépendants, ce qui
rend absurde dans ce cas le concept de distance.
On n’estime une distance qu’après avoir montré qu’il y a une liaison génétique et,
donc, une liaison physique !
Exercice 4.2
:
On étudie la ségrégation d’un couple d’allèles, notés A et a, d’un gène de
la chaîne de biosynthèse du tryptophane, chez l’ascomycète Neurospora
crassa, dont les asques sont constitués d’octades ordonnées et orientées
(avec haut et bas).
Ce gène est localisé à 5 urp (unités de recombinaison par postréduction) de
son centromère. On vous demande de décrire tous les types d’asques
possibles et d’en préciser les fréquences respectives.
Maîtriser les mécanismes de la méiose et leurs conséquences génétiques par la
reconstruction de tous les scénarios ayant conduit à une situation donnée, prendre
en compte les spécificités de Neurospora crassa.
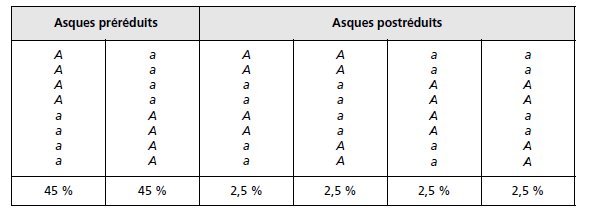
Solution. On peut observer (tabl. 4.4) deux types d’asques préréduits équifréquents et
quatre types d’asques postréduits équifréquents.
La fréquence cumulée de ces derniers a
permis d’estimer la fréquence de postréduction p qui a alors permis d’estimer la distance
du gène étudié au centromère comme égale à 100 × p/2.
Comme on nous donne cette
distance (5 urp), on peut en déduire la valeur de p, soit p = 0,1. D’où les observations
attendues.
TABLEAU 4.4.
Exercice 4.3
:
Le genre Serrospora virtualis est un genre virtuel d’ascomycètes donnant
des asques à quatre spores qui peuvent être partiellement ordonnées !
En effet les asques sont inordonnés mais on peut, en raison de la forme des
spores, distinguer les spores issues des deux chromatides « internes » et
celles issues des deux chromatides externes parce que la scission des
cellules, à la méiose I, puis à la méiose II, a laissé une marque visible sur
la paroi.
Ainsi deux des spores présentent une seule marque alors que les
deux autres spores présentent deux marques opposées.
1. Quelles sont les spores issues des deux chromatides « internes » et
celles issues des deux chromatides externes ?
2. On étudie l’aspect lisse ou rugueux de ces spores en croisant une
souche SSR à spores lisses par une souche mutante à spores rugueuses.
Interpréter les résultats (tabl. 4.5) en précisant, quand c’est possible, les
chromatides impliqués dans les crossing-over, et on calculera la distance
au centromère.
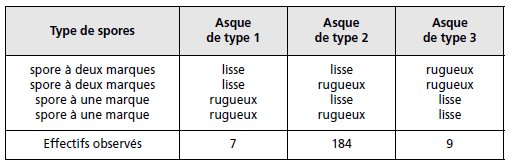
TABLEAU 4.5.
Maîtriser les mécanismes de la méiose et leurs conséquences génétiques par la
reconstruction de tous les scénario ayant conduit à une situation donnée, sous des
contraintes relatives à la perception de leur orientation spatiale.
Calcul de distance
au centromère.
Solution
1. Les spores avec deux marques correspondent aux cellules porteuses des chromatides
internes puisqu’elles portent une marque issue de la division de la méiose I, suivant le
premier plan métaphasique, puis la marque issue de la méiose II, suivant le deuxième plan
métaphasique. Les cellules porteuses des chromatides externes ne portent que la marque
issue de la méiose II.
2. Analyse de ségrégation.
Il y a ségrégation 2/2, chaque asque contenant deux spores lisses
et deux spores rugueuses. On peut considérer que les deux souches diffèrent pour un seul
gène, pour un seul des gènes impliqués dans l’aspect de la spore (voir remarque 1, ex. 4.1).
Analyse de la pré et de la postréduction.
Les asques de types 1 et 3 sont obligatoirement postréduits puisque les produits de la méiose I, répartis dans une spore uni-marquée et une
spore bi-marquée, sont toujours différents, l’une lisse et l’autre rugueuse. Ce résultat est
obtenu lorsqu’un crossing-over survient entre les chromatides 1 et 3 ou 2 et 4.
Les asques de type 2 sont un mélange d’asques préréduits et d’asques postréduits puisque les
produits de la méiose I, répartis dans une paire de spores uni-marquée et bi-marquée peuvent
être tous les deux différents ou tous les deux identiques.
Ce résultat est obtenu en cas de postréduction
lorsqu’un crossing-over survient entre les chromatides 1 et 4 ou 2 et 3.
Comme les quatre types d’asques postréduits sont équifréquents, les asques postréduits qui
ne peuvent être distingués des asques préréduits, au sein des asques de type 2, sont de
fréquence égale à ceux qui peuvent être identifiés, les asques de type 1 et 3.
Si on note p, la
fréquence de postréduction, alors chacun des quatre types d’asques postréduit aura pour
fréquence p/4 (tabl. 4.5), et les asques de type 2 auront pour fréquence (1 – p), fréquence des
asques préréduits, plus deux fois p/4, fréquences des deux types d’asques postréduits
confondus avec les préréduits.
L’identification des asques postréduits sans ambiguïté permet d’estimer la fréquence de
postréduction sachant qu’ils ne représentent que la moitié de la totalité des asques postréduits,
soit : p/2 = (7 + 9)/200, d’où p = 0,16.
Cette valeur étant inférieure à 0,66 (remarque 2, ex. 4.1), on peut conclure qu’il y a liaison
génétique entre le gène étudié et son centromère et déduire l’estimation d’une distance de
celle de la fréquence de postréduction, soit d = (p/2) × 100 = 8 urp (unités de recombinaison
par postréduction).