L’analyse de tétrades et le test de l'indépendance physique :
L’analyse de tétrades, contrairement à l’analyse génétique classique, peut souvent
statuer sur l’indépendance physique de deux gènes quand on a mis en évidence leur
indépendance génétique.
Il suffit, pour le découvrir, de confronter les domaines de
variation des valeurs des fréquences des trois types d’asques, DP, DR et T, quand les
gènes sont physiquement indépendants ou physiquement liés et rappelés ci-dessous.
De la confrontation de ces deux domaines de variation, on peut voir que :
– les méioses concernant deux gènes génétiquement indépendants mais physiquement
liés, présenteront une fréquence de tétratypes toujours égale à 2/3 (voir
page 112) alors que
– les méioses concernant deux gènes physiquement indépendants présenteront une
fréquence de tétratypes inférieure ou égale à 2/3.
Dans ces méioses la fréquence des tétratypes dépend du taux de postréduction de chacun des deux gènes et n’est
égale à 2/3 que si, au moins, un des gènes est génétiquement indépendant de son
centromère (voir page 105).
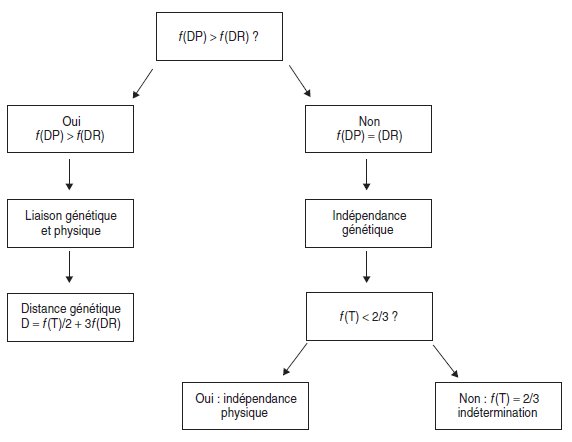
Ainsi, à la conclusion d’indépendance génétique, résultant de l’équifréquence des
DP et des DR [d’où f(GP) = f(GR)], on peut lui adjoindre, dans l’analyse de tétrades,
une conclusion d’indépendance physique si f(T) < 2/3.
Remarque 1. Dans le cas où f(T) = 2/3, la situation reste indéterminée puisque
ce cas limite peut aussi bien correspondre à une liaison physique des deux
gènes à grande distance, qu’à une indépendance physique avec indépendance
génétique entre l’un au moins de ces gènes et son centromère.
Remarque 2. Quand les gènes sont génétiquement indépendants, on a toujours
l’équifréquence des DP et des DR qui entraîne celle des gamètes parentaux et
recombinés, mais cette équifréquence toujours égale à 1/6, si les gènes sont
physiquement liés, peut être supérieure à 1/6, jusqu’à 1/2, s’ils sont physiquement
indépendants.
On définit ainsi classiquement l’organigramme décisionnel d’une analyse de
tétrades (fig. 4.10), qui ne saurait être une recette dispensant d’avoir compris tous les
phénomènes qui lui sont sous-jacents !
La
conversion génique :
A - Mise en évidence du phénomène
:
L’analyse génétique intensive de certains organismes, comme la drosophile et la
levure, a fourni des résultats assez difficiles à interpréter, comme l’illustre l’exemple
du croisement entre deux souches de Neurospora crassa à spores noires ou blanches.
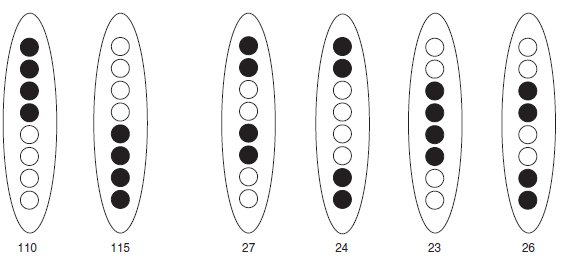
On observe 325 asques se répartissant en six types différents (fig. 4.11).
Figure 4.11
Effectifs observés de tétrades pré et postréduites à l’issue d’un croisement.
La ségrégation 2/2 observée pour la couleur de la spore permet de conclure que
les deux souches semblent différer pour un seul gène des gènes (s’ils sont plusieurs)
impliqués dans la coloration de la spore.
Par ailleurs, après avoir vérifié (valeur du
χ2 = 5,11 pour 4 ddl) une nouvelle fois l’équifréquence des deux asques préréduits,
d’une part, et l’équifréquence des quatre asques postréduits, d’autre part, on peut
calculer la fréquence de postréduction (27 + 24 + 23 + 26)/325 = 0,308, ce qui
permet d’estimer la distance du locus du gène à son centromère comme la moitié de
la fréquence de postréduction multipliée par 100, soit 15,4 unités de recombinaison
par postréduction.
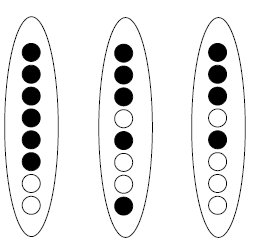
Cependant cette analyse a délaissé trois tétrades ordonnées atypiques (fig. 4.12).
Ces trois tétrades bien que minoritaires (3/325 = 0,9 %) ne peuvent pas être interprétées
par des mécanismes génétiques comme la recombinaison entre deux gènes
très liés, encore moins des mutations de novo.
• La recombinaison.
L’hypothèse de deux gènes liés avec une recombinaison très
rare entre leur locus est compatible avec la première observation, en supposant que
les spores recombinées sont de phénotype noir et qu’un crossing-over est survenu
entre les chromatides 2 et 3.
Mais il est impossible d’expliquer avec une telle hypothèse
les deux autres observations, notamment le deuxième asque où il y a cinq
spores noires et trois blanches.
En effet, si les spores recombinées sont noires, on attend six noires, si elles sont blanches on n’attend plus que deux spores noires, et si
l’une des recombinées est blanche alors que l’autre est noire, on attend quatre blanches
et quatre noires, mais jamais cinq et trois.
Figure 4.12.
Quant à la troisième tétrade, où on a bien quatre noires et quatre blanches, elle
pose un problème délicat, également posé dans l’asque précédent.
Comme la mitose
additionnelle double le nombre de cellules haploïdes, comment peut-on imaginer
qu’on puisse obtenir une spore noire de génotype (A) et une spore blanche de génotype
(a) à partir d’une spore haploïde originelle qui ne peut être à la fois (A) et (a)
puisqu’elle est haploïde ?
• La mutation. C’est un phénomène difficile à invoquer pour plusieurs raisons :
– d’une part les asques contenant des spores « mutées » semblent bien trop
fréquents (près de 1 %) pour envisager un phénomène connu pour avoir une
fréquence comprise entre 10–5 et 10–10;
– d’autre part, en supposant que le phénomène de mutation soit exceptionnellement
important pour ce gène, on devrait observer assez souvent des spores blanches
dans les croisements entre souches à spores noires ce qui n’est pas le cas;
– enfin, cette hypothèse n’est de toute façon pas cohérente avec les observations.
En
effet, la première tétrade serait facilement explicable par l’effet d’une mutation
transformant l’allèle a en A lors de la méiose I, conduisant à trois couples de
spores noires et un couple de blanches.
Mais la deuxième tétrade supposerait,
pour expliquer les trois couples de spores hétérogènes (une blanche + une noire)
que soient survenues trois mutations indépendantes, durant la duplication de
l’ADN lors de la mitose additionnelle, une de A vers a et deux de a vers A, ce qui
est totalement impensable.
La troisième tétrade supposerait deux mutations indépendantes,
ce qui est aussi trop improbable pour être cohérent avec la fréquence
observée de ces asques « aberrants ».
Puisque visiblement l’information génétique A peut se transformer en un allèle a
et réciproquement, et que ce phénomène ne saurait être une mutation, on lui a donné le nom de conversion génique; il trouve son explication dans le mécanisme moléculaire
opérant lors de certains crossing-over.
Des études récentes ont montré que, sauf
exceptions, toutes les paires de chromatides appariées à la méiose subissaient obligatoirement
au moins un crossing-over, événement indispensable à la disjonction
des paires de chromatides en anaphase 1.
Il semble cependant qu’il existe deux types
de crossing-over, les uns résultant d’une double cassure d’ADN sur deux chromatides
homologues et ceux résultant d’une simple cassure; seul ce dernier est évoqué
ci-dessous, en relation avec le modèle de Holliday.
Remarque 1. Dans les asques à spores inordonnées, la conversion génique se
traduit uniquement par un défaut de ségrégation 2/2, et chez les organismes où
seule l’analyse des gamètes en vrac est possible, elle se traduit par la survenue
de gamètes « convertis » trop fréquents pour être « mutés ».
Remarque 2. Dans la moitié des cas les gamètes « convertis » sont aussi des
gamètes recombinés par crossing-over pour des gènes localisés de part et
d’autre du gène ayant subi la conversion, ce qui a amené les chercheurs à
supposer que la conversion était associée au mécanisme du crossing-over.
B - Interprétation moléculaire de la conversion génique
:
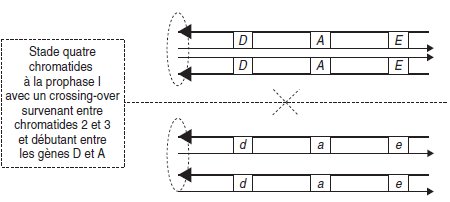
Il est nécessaire (fig. 4.13) de considérer, par un trait d’épaisseur différente, les deux
brins complémentaires de chaque molécule d’ADN, ainsi que leur orientation 5′ vers
3′(sens de la flèche).
Figure 4.13.
Dans la mesure où le crossing-over n’intéresse que les chromatides 2 et 3, seules
ces chromatides seront suivies, les deux autres donnant des gamètes parentaux pour
les trois gènes.
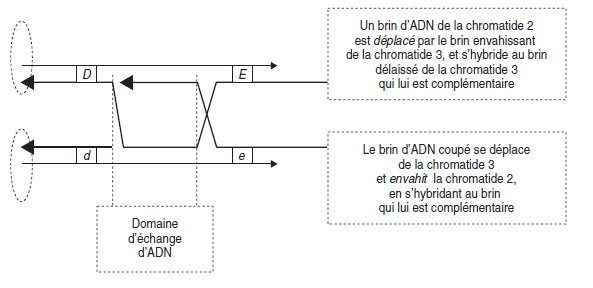
Un modèle théorique du crossing-over consiste à supposer qu’il est initié par la
coupure d’un brin d’ADN (par exemple ici sur la chromatide
3) qui peut alors se déshybrider de son brin complémentaire pour envahir le duplex homologue de la
chromatide 2, en s’hybridant au brin qui lui est complémentaire, et en repoussant son homologue de la chromatide 2 vers le brin complémentaire non coupé de la chromatide
3, donneuse du brin envahissant (fig. 4.14).
Figure 4.14.
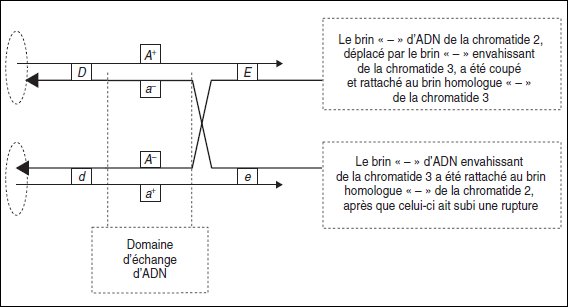
Le brin de la chromatide 3 envahissant la chromatide 2 est alors rattaché à l’extrémité
5′ libre de celle-ci, tandis que le brin de la chromatide 2, déplacé par le brin
envahissant, est rompu puis rattaché à l’extrémité 5′ libre de la chromatide 3
(fig. 4.15).
Le résultat revient à considérer que tout s’est passé comme si deux brins d’ADN
homologues avaient été coupés en même temps, au même site, puis échangés et
rattachés, mécanisme initialement imaginé par Holliday. Le mécanisme de CO issu
d’une double coupure ne sera pas envisagé ici.
Figure 4.15.
Dans la structure moléculaire ainsi obtenue (fig. 4.15) les deux chromatides ne
sont plus indépendantes puisqu’elles ont échangé un de leur brin d’ADN, mais un
seul (ici les brins « – ») sur une certaine zone.
La structure formée par les deux chromatides
et leur quatre brins est appelée structure en χ de Holliday.
Remarque. Il est important de noter que les informations génétiques codées
par les deux brins échangés sont A– et a–, dans leur séquence nucléotidique de
type brin « – », ce qui entraîne obligatoirement, quelque part dans la zone
d’échange, un misappariement puisque les brins a+ et A+ auxquels ils sont
respectivement appariés (fig. 4.15) ne leurs sont pas strictement complémentaires.
a) Résolution de la structure en χ de Holliday
:
L’échange des deux brins « – » d’ADN se réalise sur une certaine distance.
La poursuite
de la division nucléaire de la méiose I impose que la structure de Holliday soit
« résolue » en deux chromatides indépendantes qui pourront migrer vers chacun des
pôles.
L’analyse des tétrades ou des octades ordonnées a montré que deux résolutions
différentes et équiprobables pouvaient survenir.
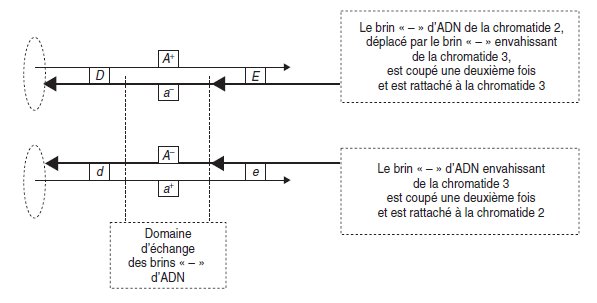
Figure 4.16.
• Le premier type de résolution (fig. 4.16) est un arrêt de l’échange par double
rupture des deux brins échangés, sous l’action d’une topo-isomérase, suivie de
l’échange en sens inverse et rattachement aux chromatides d’origine; cette résolution
n’entraîne aucune recombinaison génétique de part et d’autre de l’échange mais
laisse, dans la zone d’échange, une molécule d’ADN hétéroduplex (voir plus bas).
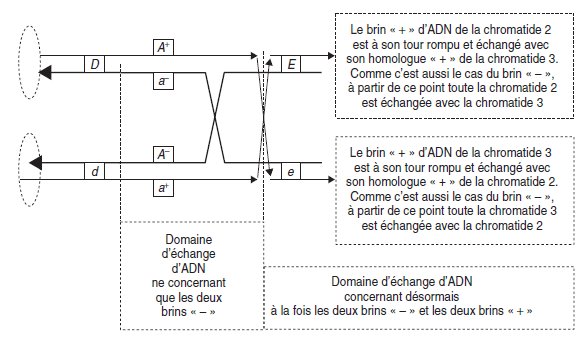
• Le deuxième type de résolution (fig. 4.17) est une poursuite de l’échange initial
(ici les brins « – ») et l’initiation d’un échange sur les deux brins non concernés par le premier échange (ici les brins « + »).
Il s’agit alors du crossing-over stricto sensu,
avec un véritable échange chromatidique concernant les deux brins d’ADN, ce qui
conduit, de part et d’autre des points d’initiation des deux échanges, à une recombinaison
génétique, et laisse entre ces deux points une zone d’ADN hétéroduplex,
comme dans le cas précédent.
Figure 4.17.
b) Conversion génique résultant de la réparation de la zone hétéroduplex
:
Il est très important de noter que sur toute la zone d’échange simple brin entre les
deux chromatides, du point d’initiation au point de résolution de la structure de
Holliday, la molécule d’ADN résultante constitue un hétéroduplex, avec un brin « + »
d’une chromatide et un brin « – » d’une autre chromatide.
La conséquence génétique
de la formation d’une molécule hétéroduplex est que la complémentarité des deux
brins est localement imparfaite quand il y a une variation allélique de la séquence;
c’est par exemple le cas pour le gène A où chaque molécule d’hétéroduplex est en
fait porteuse des deux informations alléliques A et a, l’une sur le brin sens, avec la
séquence A+, l’autre sur le brin antisens, avec la séquence a–, et réciproquement.
Dans cette zone hétéroduplex de la molécule d’ADN, la différence de message
génétique entre les allèles A et a du gène se traduit par une complémentarité
chimique imparfaite, par un misappariement de une ou plusieurs paires de bases,
uniquement au(x) site(s) du gène où le message génétique A diffère du message
génétique a; à ce niveau les brins d’ADN porteurs des séquences A+ et a–, ou entre
a+ et A–, ne peuvent être parfaitement appariés.
Ce misappariement peut être détecté par la cellule, ce qui conduit à sa « réparation
»; l’un des deux brins est clivé de part et d’autre du misappariement, puis la lacune est comblée par une ADN-polymérase qui utilise l’autre brin comme matrice.
Ainsi l’allèle A peut être « totalement reconstitué » sur les deux brins, si le brin hétéroduplex
portant a– (ou a+) est clivé, et l’allèle a « totalement reconstitué » si c’est le
brin de l’hétéroduplex portant A+ (ou A–) qui est clivé.
Comme les deux réparations sur les deux hétéroduplex des chromatides 2 et 3
sont indépendantes, la correction peut parfaitement se faire dans le même sens sur
les deux chromatides, ce qui aboutira, par exemple, à deux allèles A, et donc à une
méiose conduisant à trois allèles A pour un seul a, c’est-à-dire à la conversion
génique d’un allèle a en un allèle A.
Il est ainsi possible d’expliquer les asques avec six spores noires et deux blanches,
la mitose additionnelle n’ayant fait que doubler le nombre de spores.
La conversion génique est donc un phénomène qui touche les gènes compris dans
la zone d’échange d’un brin d’ADN au cours d’un crossing-over si ces gènes présentent
une variation allélique.
La formation dans cette zone d’une molécule d’ADN hétéroduplex, porteuse de deux séquences alléliques différentes conduit à un misappariement
local au site de la variation allélique du gène, là où les deux brins ne sont
plus strictement complémentaires.
La réparation du misappariement par excision-resynthèse d’un des deux brins,
désigné aléatoirement, conduit à la reconstitution d’un homoduplex (deux brins
strictement complémentaires) porteur d’un seul message allélique.
Les deux chromatides
peuvent voir leurs hétéroduplex corrigés dans le même sens, vers le même
allèle, de sorte que la situation d’hétérozygotie qui prévalait avant le « passage » du
crossing-over est remplacée par une homozygotie en faveur d’un des deux allèles;
l’un des deux allèles a été « converti » en l’autre.
Bien évidemment, une cellule peut ne pas avoir le temps de réparer les hétéroduplex,
surtout quand elle entre presque immédiatement dans une nouvelle division, ce
qui est le cas chez les ascomycètes avec la mitose additionnelle.
Lors de la phase de
réplication, la zone hétéroduplex donnera deux homoduplex génétiquement différents,
l’un porteur de l’allèle A, à partir du brin A+ (ou A–) utilisé comme matrice,
l’autre porteur de l’allèle a, à partir du brin a– (ou a+) utilisé comme matrice.
Ce qui
explique nos octades où une cellule haploïde a pu donner une spore noire et une
spore blanche, car, au niveau du gène concerné, sa molécule d’ADN était constituée
d’un hétéroduplex porteur simultanément (mais de manière transitoire et non
complémentaire) des deux informations alléliques.