B - Analyse de tétrades pour deux gènes physiquement liés :
a) Les configurations possibles de la méiose et des asques résultants
:
Si deux gènes sont physiquement liés, les configurations possibles de la méiose
dépendront d’abord de la disposition aléatoire des deux centromères de la paire
d’homologues, ensuite de la survenue ou non d’un ou plusieurs crossing-over, enfin,
dans le cas de plusieurs crossing-over, des chromatides impliquées.
Comme, ici, la distinction entre haut et bas de l’asque double très exactement
chacune des configurations (ce qui n’est pas tout à fait le cas pour deux gènes physiquement
indépendants où on passe de 20 à 36 configurations), nous ne ferons pas
cette distinction.
Notre intérêt étant essentiellement centré sur les événements survenant entre les
locus des deux gènes, nous négligerons la position du centromère qui ne sera figuré
en pointillés que pour rappeler qu’il unit en un site les deux chromatides soeurs.
Dans ces conditions, le croisement d’une souche haploïde {D; E} par une souche
haploïde {d; e}, sachant que les gènes D et E sont physiquement liés, donne une
cellule diploïde {AD//de} dont la méiose présente plusieurs configurations possibles
(fig. 4.6 à 4.8) :
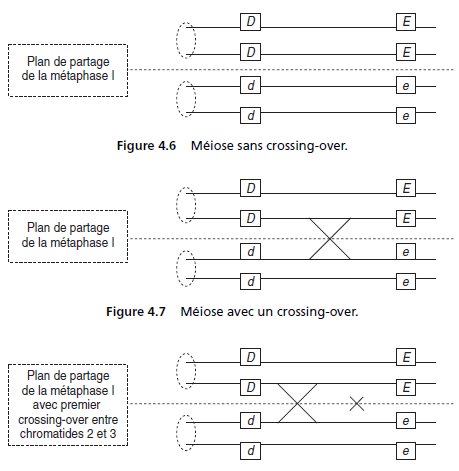
• Cas 1, il n’y a aucun crossing-over entre les locus des deux gènes (fig. 4.6) : la
méiose donne exclusivement un ditype parental (DP) avec deux gamètes de type
parental {D; E} et deux gamètes de type parental {d; e}.
• Cas 2, il y a un crossing-over entre les locus des deux gènes (fig. 4.7) : la méiose
donne exclusivement un tétratype (T) avec un gamète de type parental {D; E}, un
gamète de type parental {d; e}, un gamète de type recombiné {D; e} et un gamète de
type recombiné {d; E}, quelle que soit la paire de chromatide impliquée par le
crossing-over.
Remarque. Dans un asque ordonné, les positions respectives des spores
parentales et recombinées dépendent des chromatides impliqués par le
crossing-over (1-4, ou 1-3 ou 2-4 ou 2-3).
Le fait que les quatre types de
dispositions soient équifréquentes est une nouvelle validation de l’hypothèse
selon laquelle les chromatides sont aléatoirement impliquées par un crossingover.
• Cas 3, il y a deux crossing-over entre les locus des deux gènes (fig. 4.8) : si le
premier crossing-over survient entre les chromatides 2-3, le type d’asque résultant de la méiose dépendra des chromatides impliquées dans le deuxième crossing-over.
Il est aisé de voir qu’on obtiendra :
– un DP si le deuxième crossing-over touche les mêmes chromatides 2-3;
– un DR si le deuxième crossing-over touche les deux autres chromatides 1-4;
– un T si le deuxième crossing-over touche une même chromatide, 2 ou 3, et une
troisième chromatide non soeur, c’est-à-dire si le deuxième crossing-over est 2-4
ou 3-1.
Ainsi, dans le cas d’un double crossing-over entre deux locus, on obtient statistiquement
1/4 de DP + 1/2 de T + 1/4 de DR.
• Cas 4, il y a plus de deux crossing-over entre les locus des deux gènes; on montre
alors que les DP et les DR sont, là encore, équifréquents.
Figure 4.8 Méiose avec deux crossing-over.
b) Fréquences des tétrades ou des gamètes
:
La fréquence de crossing-over entre deux locus est une fonction de la distance entre
ces locus.
Aussi les fréquences des trois types de tétrades et celles des gamètes
parentaux (GP) ou recombinés (GR) le sont également. Il est donc facile de tirer les
fréquences gamétiques des fréquences des types d’asques :
• Si la distance est très faible, voire nulle, entre les deux locus, il n’y aura jamais de
crossing-over, et les méioses ne donneront que des DP (cas 1).
On aura :
Ce cas correspond à la liaison génétique et physique absolue.
• Si la distance est assez grande pour qu’un ou plusieurs crossing-over puissent
survenir (cas 2, 3 ou 4) mais qu’elle est encore suffisamment peu élevée pour qu’une
fraction des méioses se déroule sans crossing-over (cas 1), cette fraction donnera un
surplus de DP et donc de GP, puisque les méioses avec crossing-over donnent statistiquement
autant de DP que de DR, donc autant de GP que de GR.
On aura :
Ce cas correspond à celui déjà vu dans l’analyse génétique classique où la
fréquence des gamètes recombinés étant inférieure à celle des gamètes parentaux, on
peut conclure à la liaison génétique, à la liaison physique, et calculer une distance en
unités de recombinaison génétique, égale à f(GR) × 100.
Cette distance peut aussi s’écrire :
qui n’est qu’une façon différente d’écrire la distance en fonction de la fréquence des
gamètes recombinés.
Remarque. On verra, un peu plus loin, que l’analyse des tétrades permet de
définir une distance beaucoup moins sous-estimée que la distance en unités de
recombinaison génétique.
• Si la distance est assez grande pour qu’au moins un crossing-over survienne dans
chaque méiose, aucune méiose ne se déroulera sans crossing-over (cas 1 devenu
inexistant), il n’y aura donc pas surplus de DP ni de GP, puisque les méioses avec
crossing-over donnent statistiquement autant de DP que de DR, donc autant de GP
que de GR.
On aura :
Ce cas correspond à celui déjà vu dans l’analyse génétique classique où la
fréquence des gamètes recombinés étant égale à celle des gamètes parentaux, on
peut conclure à l’indépendance génétique.
C - Domaine de variation des trois types de tétrades
pour deux gènes physiquement liés :
Deux gènes physiquement liés sont génétiquement liés quand leur distance est telle
qu’une fraction des méioses se déroule sans crossing-over entre leurs locus, ce qui
entraîne un surplus de DP, et donc de gamètes parentaux (voir ci-dessus).
Deux gènes physiquement liés sont génétiquement indépendants quand leur
distance est telle qu’au moins un crossing-over survienne dans chaque méiose, ce
qui conduit à l’égalité des DP et des DR, et à l’équifréquence des gamètes parentaux
et des gamètes recombinés (voir ci-dessus).
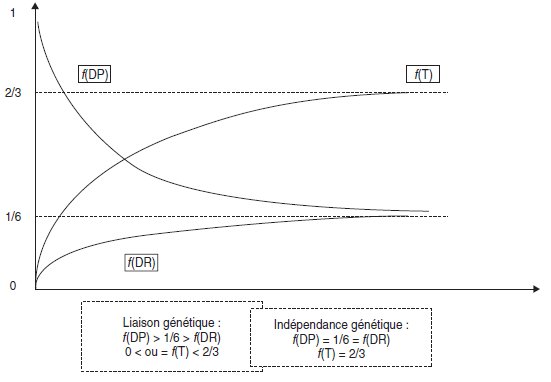
Dans ce cas limite d’indépendance génétique, les fréquences des DP, DR et T
prennent des valeurs limites égales à 1/6, 1/6 et 2/3.
En effet, si les gènes sont indépendants, les deux paires d’allèles ségrègent indépendamment.
• Le premier des deux allèles D peut ségréger à la méiose I avec un allèle E, événement
de probabilité égale à 1/2 (il y a quatre allèles disponibles), le deuxième allèle D
ségrégera :
– soit avec un allèle E, événement de probabilité 1/3 (il n’y a plus que trois allèles
disponibles dont un seul allèle E), ce qui donnera des DP, car les deux premiers
gamètes étant {D; E}, les deux autres sont obligatoirement {d; e};
– soit avec un allèle e, événement de probabilité 2/3 (il n’y a plus que trois allèles
disponibles mais deux sont e), ce qui donnera des T, car les deux premiers gamètes
étant {D; E} et {D; e}, les deux autres sont obligatoirement {d; E} et {d; e}.
• Le premier des deux allèles D peut ségréger à la méiose I avec un allèle e, événement
de probabilité égale à 1/2 (il y a quatre allèles disponibles), le deuxième allèle D
ségrégera :
– soit avec un allèle e, événement de probabilité 1/3 (il n’y a plus que trois allèles
disponibles dont un seul allèle e), ce qui donnera des DR, car les deux premiers
gamètes étant {D; e}, les deux autres sont obligatoirement {d; E};
– soit avec un allèle E, événement de probabilité 2/3 (il n’y a plus que trois allèles
disponibles mais deux sont E), ce qui donnera des T, car les deux premiers gamètes
étant {D; e} et {D; E}, les deux autres sont obligatoirement {d; E} et {d; e}.
En sommant toutes ces probabilités, on voit bien qu’à l’issue d’une série de
méioses pour deux gènes physiquement liés, mais génétiquement indépendants, on
aura :
Le domaine de variation des fréquences des trois types d’asques est précisé par un
graphe entre leurs valeurs limites correspondant à la liaison absolue, d’une part, et
l’indépendance génétique, d’autre part (fig. 4.9).
Figure 4.9.
C - L’analyse de tétrades et la correction de la distance génétique
:
La fréquence des crossing-over entre deux locus étant une fonction de leur distance,
on peut estimer cette distance par l’estimation de la fréquence des crossing-over, et
celle-ci ne sera accessible qu’à travers la fréquence des objets générés par ces
crossing-over.
Dans l’analyse génétique classique, ces objets sont les gamètes recombinés, mais
la fréquence des gamètes recombinés n’est un reflet exact de celle des crossing-over
que si la distance est suffisamment faible pour qu’il n’y ait qu’un crossing-over au
plus entre les deux gènes.
La distance génétique en unités de recombinaison est donc correctement estimée
pour les petites distances mais devient vite biaisée pour les grandes distances, car la
survenue de plusieurs crossing-over entre les deux gènes peut reconstituer des
gamètes parentaux, ce qui entraîne une sous-estimation des événements de recombinaison
par crossing-over et, par conséquent, de la distance.
Haldane a montré qu’on pouvait, par une transformation mathématique, définir
une métrique additive fonction de la fréquence de gamètes recombinés.
L’analyse de tétrades permet aussi la définition d’une distance corrigée, moins
biaisée que la distance en unités de recombinaison, parce qu’on peut estimer directement
la fréquence d’un autre événement généré par les crossing-over, celle des chromatides
remaniés, que les gamètes soient parentaux ou recombinés.
En effet, il suffit de se rappeler que l’absence de crossing-over aboutit exclusivement
à des DP, que la survenue d’un seul crossing-over donne exclusivement des T,
tandis que la survenue de deux crossing-over fournit 1/4 de DP + 1/2 de T + 1/4 de DR.
On peut ainsi évaluer la fréquence des méioses impliquant, entre chacun des deux
gènes, zéro, un ou deux crossing-over.
• La fréquence des méioses avec zéro crossing-over entre les deux gènes est égale à
la fréquence des DP, à condition de retirer les DP obtenus avec deux crossing-over.
Or celle-ci est connue, elle est égale à la fréquence des DR, puisque les DP issus de
deux crossing-over sont équifréquents aux DR.
Ainsi :
f(méioses avec zéro crossing-over) = f(DP) – f(DR)
• La fréquence des méioses avec un seul crossing-over entre les deux gènes est égale
à la fréquence des T, à condition de retirer les T obtenus avec deux crossing-over.
Or
celle-ci est connue, elle est égale au double de la fréquence des DR, puisque les T
issus de deux crossing-over sont deux fois plus fréquents que les DR.
Ainsi :
f(méioses avec un crossing-over) = f(T) – 2f(DR)
• La fréquence des méioses avec deux crossing-over entre les deux gènes est égale à
quatre fois la fréquence des DR, puisque la survenue de deux crossing-over ne donne
des DR qu’une fois sur quatre.
Ainsi :
f(méioses avec deux crossing-over) = 4f(DR)
Il s’agit maintenant d’estimer la fréquence des chromatides remaniées :
– quand il y a zéro crossing-over, aucune chromatide n’est remaniée;
– quand il y a un crossing-over, deux des quatre chromatides sont remaniées, seule
la moitié des chromatides est remaniée;
– quand il y a deux crossing-over, les quatre chromatides sont remaniées, comme
dans les DR.
Chez les DP, deux chromatides sont remaniées deux fois, ce qui revient au même
que quatre chromatides remaniées une fois.
Chez les T, deux chromatides sont remaniée
une fois et une chromatide est remaniée deux fois, ce qui revient au même que
quatre chromatides remaniées une fois.
On peut donc considérer que les méioses avec deux crossing-over sont assimilables
à des méioses où toutes les chromatides sont remaniées une fois (ou un équivalent) tandis que les méioses avec un crossing-over sont des méioses ne renfermant
qu’une moitié de chromatides remaniées. Ainsi, la fréquence des chromatides remaniées
est égale à :
f(chr. r.) = f(méioses avec un crossing-over)/2 + f(méioses avec deux crossing-over)
ce qui aboutit à :
f(Chr.r.) = [f(T) – 2f(DR)]/2 + [4f(DR)]
soit :
qui diffère de la fréquence des gamètes recombinés qui n’est égale qu’à :
f(GR) = f(T)/2 + f(DR)
La fréquence de chromatides remaniées ainsi obtenue est plus précise, moins
sous-évaluée, que la fréquence des gamètes recombinés.
Elle est corrigée par un
terme additionnel de 2f(DR), correspondant aux DP obtenus par double crossingover,
et à la moitié des T obtenus par double crossing-over, c’est-à-dire, très exactement,
la fréquence de tous les gamètes parentaux ayant subi un crossing-over, non
recensables par l’analyse classique, mais recensables par l’analyse de tétrades.