C - Interprétation fonctionnelle et moléculaire
de la dominance et la récessivité :
Seule l’analyse moléculaire du gène ou de son produit, dans leurs diverses formes
alléliques, en rapport avec la fonction du gène et les phénotypes associés, permet
une approche fonctionnelle de l’effet des mutations et une interprétation des effets
de dominance et de récessivité.
Pour illustrer cette démarche, on prendra tous les exemples dans la pathologie
humaine où il est possible d’observer les quatre cas possibles (tableau 5.4) des mutations
de perte de fonction responsables d’un phénotype récessif ou dominant, puis des
mutations de gain de fonction, responsable d’un phénotype récessif ou dominant.
– Si la mutation est une perte de fonction (absence de produit ou produit inactif), le
phénotype muté peut être récessif, soit parce que l’effet de l’allèle sauvage chez le
diploïde hétérozygote « compense » totalement l’absence d’effet de l’allèle muté,
c’est le cas pour certaines mutations de perte de fonction concernant des gènes
dont l’expression est régulée avec précision (mutations thalassémiques dans le
gène β de l’hémoglobine); soit parce que l’effet de l’allèle sauvage fonctionnel
est haplo-suffisant et que 50 % de la quantité normale de produit suffit largement
à assurer une physiologie normale et conduit donc à un phénotype sauvage (cas de
la plupart des pertes de fonction de gènes du métabolisme).
– Si la mutation est une perte de fonction (absence de produit ou produit inactif), le
phénotype muté peut être dominant pour plusieurs causes fonctionnelles possibles.
Le phénotype muté est dominant quand l’effet de l’allèle sauvage, chez le
diploïde hétérozygote, ne peut pas « compenser » totalement l’absence d’effet de
l’allèle muté.
On dit alors qu’il y a dominance par haplo-insuffisance; cette haploinsuffisance
(il n’y a qu’un seul gène actif sur les deux) ne permet pas à la cellule,
au tissu, à l’organisme d’avoir la quantité suffisante de produit du gène pour
assurer la fonction de celui-ci; c’est le cas notamment pour les diabètes de type
MODY, ou les hypercholestérolémies familiales.
On peut d’ailleurs remarquer
que la maladie est presque toujours plus grave chez les homozygotes porteurs
d’une mutation dominante par haplo-insuffisance, par exemple les hypercholestérolémies,
ce qui est en accord avec le fait que le phénotype résulte d’un effet dose
du produit codé par le gène impliqué.
Cas particulier. Le phénotype muté peut « paraître » dominant parce que sa
transmission est associée à la transmission d’une perte de fonction récessive
mais que l’organisme va, dans le cours de son développement, perdre la copie
haplo-suffisante dans certains sous clones somatiques; ces sous clones seront
alors porteurs de deux copies non fonctionnelles du gène et à l’origine du
phénotype muté de l’organisme (cas des formes héréditaires de cancers).
Remarque. Ce dernier cas montre qu’il est abusif de considérer qu’un phénotype
dominant serait associé à une mutation dont l’effet serait dominant sur
celui de l’allèle sauvage.
– Si la mutation est un gain de fonction, un produit muté du gène est présent.
Si le
phénotype associé est récessif, il faut alors considérer que l’effet de l’allèle sauvage
(plutôt du produit de cet allèle) chez l’hétérozygote « masque » l’effet du produit
muté; c’est le cas de la mutation drépanocytaire (voir plus loin).
Remarque.
Ce dernier exemple montre qu’il est abusif de considérer qu’un
phénotype récessif serait associé à « un allèle muté qui ne s’exprimerait pas ».
Au contraire, dans cet exemple, l’allèle drépanocytaire n’est pas une perte de
fonction et il s’exprime, mais son expression n’est pas perceptible au niveau
du phénotype étudié, les effets cliniques.
– Si la mutation est un gain de fonction, un produit muté du gène est présent.
Si le
phénotype est dominant, on peut considérer que son action spécifique s’impose
face à celle du produit sauvage, notamment quand cette action est toxique sur la
cellule, le tissu ou l’organisme, et que le produit sauvage codé par l’autre allèle
n’est pas en mesure de contrecarrer l’effet du produit muté, soit pour des raisons
quantitatives, soit pour des raisons qualitatives.
Ce cas correspond à toutes les
maladies neuro-dégénératives (Huntington, ataxies spino-cérébelleuses, vraisemblablement
Alzheimer) ou la Dystrophie myotonique.
On dit alors le phénotype
muté est dominant par « effet dominant négatif » de la mutation sur l’effet de
l’allèle sauvage.
Remarque 1. Dominance ou récessivité d’un phénotype mutant ne sont donc
que statistiquement associés au fait que la mutation responsable soit un gain
ou une perte de fonction.
Si on observe fréquemment que les mutations de
perte de fonction dans certains gènes ont un effet récessif vis-à-vis de l’effet
du gène sauvage, il arrive aussi assez souvent, dans d’autres gènes, qu’elles
puissent être responsables d’un phénotype mutant « dominant » par haploinsuffisance
de la copie sauvage du gène (en fait, il y a plutôt co-dominance
car si l’hétérozygote est de phénotype mutant, l’homozygote muté peut présenter
un phénotype muté beaucoup plus marqué ou grave).
Inversement, si de
nombreuses maladies dominantes résultent de l’effet « dominant négatif »
d’un allèle muté sur l’effet de l’allèle sauvage, on ne doit pas négliger l’existence
de maladies (ou de trait) récessives associées à des mutations de gain de
fonction.
Remarque 2. Il faut rappeler que la dominance et la récessivité sont des attributs
du phénotype et que c’est par un abus de langage qu’on parle de mutation
dominante ou récessive :
➤ d’une part, quand on dit cela, on sous-entend que c’est par rapport à
l’allèle sauvage, car un génotype β 0//β S aura un phénotype drépanocytaire,
ce qui signifie que l’allèle β S qui est récessif vis-à-vis de β A, pour le
phénotype maladie, est dominant vis-à-vis de β 0;
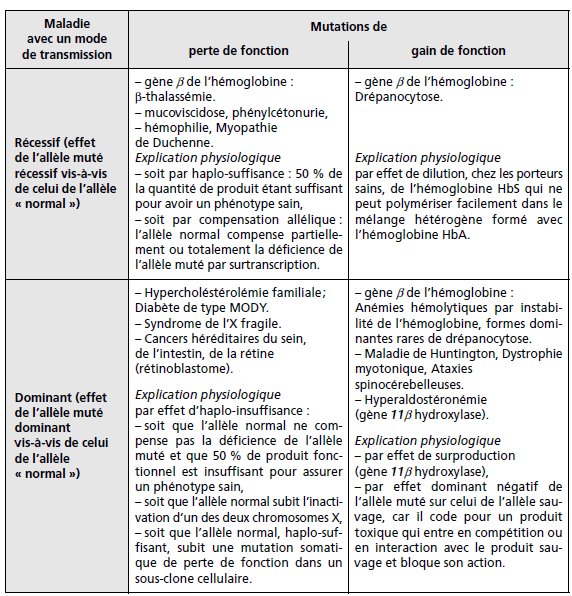
TABLEAU 5.4 EXEMPLES DES PATHOLOGIES HUMAINES.
Les mutations de perte de fonction ont le plus souvent un effet récessif vis-à-vis de
celui de l’allèle normal alors que les mutations de gain de fonction ont le plus souvent
un effet dominant, mais cette règle n’est que statistique et des maladies dominantes
peuvent être associées à des pertes de fonction alors que certaines maladies récessives
sont associées à des gains de fonction.
➤ d’autre part, quand on dit cela, on se réfère à un caractère particulier avec
un crible phénotypique, car l’analyse des mêmes allèles dans un autre
caractère peut changer leur relation de dominance. Par exemple, pour les
phénotypes électrophorétiques des dimères αβ issus de l’hémoglobine en
solution aqueuse, les trois génotypes β A//β A, β S//β S, β A//β S présentent trois
phénotypes différents, respectivement une bande rapide, une bande lente,
les deux bandes, ce qui signifie, que pour le caractère de mobilité électrophorétique,
les deux allèles sont codominants.
Exercice 5.1
:
On dispose de deux souches de levure, l’une de phénotype [mat a, ura–]
auxotrophe pour l’uracile, l’autre de phénotype [mat α, gal–], incapable de
consommer le galactose. On sait que les phénotypes mutés de ces deux
souches sont récessifs.
Question 1.
Quelle sera la composition du milieu de culture de chacun des deux mutants
et de la boîte de croisement ? Justifiez vos réponses.
Question 2.
À partir de la souche [mat a, ura–], on a obtenu une série de 10 mutants
auxotrophes pour l’histidine, nommés m1 à m10.
L’étude de mutants m1 à
m8 a montré qu’il s’agissait de mutants récessifs, différant de la souche
sauvage pour un seul gène.
Comment s’y est-on pris pour démontrer cela avec le matériel dont nous
disposons ?
Il convient donc de définir le protocole expérimental suivi (les croisements,
les milieux des boîtes de croisement ou de recueil des spores, les boîtes de
repiquage) et de préciser, à chaque étape les observations, qualitatives et/
ou quantitatives, qui justifient les conclusions rapportées.
Question 3.
L’analyse du mutant m9 a montré qu’il s’agissait d’un mutant récessif
affecté dans deux gènes indépendants.
Précisez, sans le reprendre point par
point, à quelle(s) étape(s) du protocole de la question 2, une (ou des) observation(
s) particulière(s) permettent de conclure pour ce mutant m9.
Question 4.
L’analyse du mutant m10 a montré qu’il s’agissait d’un mutant dominant
affecté dans un seul gène.
Précisez, sans le reprendre point par point, à
quelle(s) étape(s) du protocole de la question 2, une (ou des) observation(s)
particulière(s) permettent de conclure pour ce mutant m10.
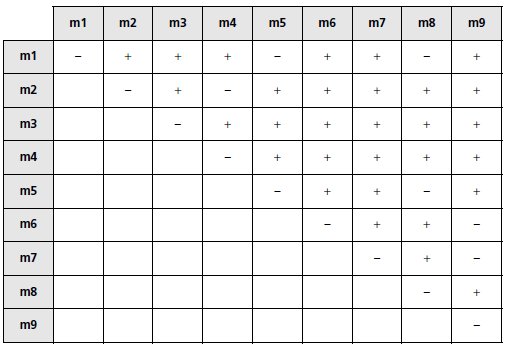
Question 5.
On réalise les croisements entre mutants en utilisant des spores issues des
croisements précédents et porteuses du signe sexuel adéquat ainsi que des
marqueurs de sélection des diploïdes nécessaires. On obtient les résultats
suivants :
Quelles sont les conclusions génétiques qui découlent de l’analyse de ce
tableau ? Sont-elles cohérentes avec les informations recueillies auparavant ?
– Maîtriser la réalisation et l’interprétation du test de complémentation fonctionnelle.
– Rapporter ses résultats à ceux acquis dans l’analyse de la ségrégation.
Solution
1. [mat a, ura–] sur Mo + ura; [mat α, gal–] sur Mo; les diploïdes sur Mo(gal)
2. a. Le mutant mi est croisé avec [mat a, gal–, his+], sauvage pour le caractère histidine, sur
une boîte Mo(gal) + his, car on ne sait si le mutant est récessif ou dominant : on obtient des
colonies de diploïdes.
b. les colonies sont répliquées sur Mo et y poussent ce qui prouve que les diploïdes sont
[his+] et que les mutants sont récessifs.
c. On conduit les diploïdes à la méiose et on recueille les spores sur un milieu Mo + ura + his,
on teste les colonies sur un milieu Mo + ura pour dénombrer les colonies [his+] et avoir par
différence le dénombrement sur la boîte mère des colonies [his–] : on observe 50 % de chaque
type, ce qui indique que chaque mutant diffère de la souche sauvage pour un seul gène relativement
au caractère histidine, un seul des gènes de la chaîne de biosynthèse de l’histidine.
3. À l’étape c, on n’observe pas 50 % de [his+], mais 25 %, et pas 50 % de [his–] mais 75 %.
4. À l’étape b, on n’obtient pas de colonies sur la boîte de réplique, ce qui prouve que les
diploïdes, hétérozygotes, sont [his–].
5. Les groupes de complémentation permettent de définir 5 gènes et le mutant m9, muté dans
deux gènes apparaît bien dans deux groupes.
Ces groupes sont : (1, 5, 8) (2, 4) (3) (6, 9) (7, 9)