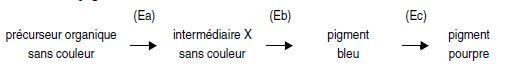Chez Lathyrus odoratus, on connaît trois enzymes Ea, Eb et Ec, respectivement
produits des gènes A, B et C, qui gouvernent la synthèse de l’antocyane,
le pigment pourpre des pétales, selon la réaction (on sait qu’en
absence de pigments, les fleurs sont blanches) :
1. Quelle sera la couleur des fleurs dans une souche pure dépourvue d’une
des trois activités ? (Les trois souches pures sont appelées a, b et c).
2. On croise chacune des souches a, b ou c avec la souche pure sauvage, on
observe des descendants F1 aux fleurs pourpres. Interprétez.
3. On croise chacune des souches mutantes a, b et c entre elles.
Quelle sera
la couleur des fleurs en F1 ?
4. On découvre une nouvelle variété aux fleurs blanches, appelée d qui,
croisée avec la souche sauvage ou les souches a, b ou c, donne des F1 aux
fleurs pourpres. Concluez.
5. Aurait-on pu répondre à la question précédente en l’absence du résultat
du croisement entre la souche d et la souche sauvage ?
6. Il est facile d’injecter, à l’ouverture du bourgeon floral, un extrait purifié
du pigment bleu ou de l’intermédiaire X sans couleur.
a. Quelle serait la couleur des fleurs chez un individu c, ayant reçu un
extrait de pigment bleu; de produit X ?
b. Quelle serait la couleur des fleurs chez un individu b, ayant reçu un
extrait de pigment bleu; de produit X ?
c. On injecte le produit X dans le bourgeon floral d’un individu d, les fleurs
sont blanches. Concluez.
– Maîtriser l’interprétation fonctionnelle de la compensation allélique et de la
complémentation fonctionnelle.
– Comprendre l’effet de la position d’un gène (de son produit) dans une chaîne
sur le phénotype.
Solution
1. • Souche pure a. Son génotype est a–//a–; absence de l’activité Ea : fleurs blanches; accumulation
du précurseur organique;
• souche pure b. Son génotype est b–//b–; absence de l’activité Eb : fleurs blanches; accumulation
de l’intermédiaire X, sans couleur;
• souche pure c. Son génotype est c–//c–; absence de l’activité Ec : fleurs bleues; accumulation
du précurseur constituant un pigment bleu ne pouvant être transformé en antocyane
pourpre.
2. Les trois phénotypes mutants sont récessifs; l’effet des mutations est récessif face à celui
de l’allèle sauvage qui compense l’effet (ou l’absence d’effet) de l’allèle muté.
3. Connaissant l’effet récessif des mutations et sachant que les trois mutations étudiées
touchent des gènes différents, les diploïdes issus du croisement entre deux souches quelconques
présenteront une complémentation fonctionnelle conduisant à un phénotype sauvage
[fleurs pourpres].
4. La souche d présente un phénotype récessif, ce qui permet d’interpréter les résultats des
croisements entre mutants, du point de vue fonctionnel.
Dans tous les cas, il y a complémentation
fonctionnelle, puisque le diploïde issu du croisement entre d et l’un des trois autres
mutants, a, b ou c, présente un phénotype sauvage [fleurs pourpres].
La souche d est donc
mutée dans un autre gène que les gènes A, B ou C; il existe donc au moins quatre gènes dans
la chaîne de biosynthèse de l’antocyane pourpre.
5. Le test de dominance/récessivité des mutants est nécessaire pour pouvoir interpréter
l’absence de complémentation et conclure que deux mutations touchent le même gène.
Dans le cas, comme celui présenté ici, où il y a complémentation fonctionnelle, les mutants
sont obligatoirement récessifs, et le test de dominance ne fait que confirmer cet état sans être
absolument obligatoire à l’interprétation du croisement entre mutants.
En d’autres termes, on
aurait pu aussi bien répondre à la question 5 sans avoir le résultat du croisement entre la
souche d et la SSR de la question 4 !
6. a. Aucun effet, la souche ne peut transformer le pigment bleu.
b. Fleur rouge, quand on apporte le pigment bleu que la souche b ne peut produire par
absence de l’activité Eb.
Fleur blanche si on apporte X, puisque X ne peut être transformé.
c. Si, en injectant X, les fleurs demeurent blanches, c’est que X ne peut être transformé en antocyane pourpre et que la chaîne de biosynthèse se trouve bloquée avant la formation du
pigment bleu.
Le gène d code pour une enzyme gouvernant une étape située entre X et le
pigment bleu ou entre le précurseur organique et X.
On peut aussi imaginer qu’il existe entre
le pigment bleu et l’antocyane un intermédiaire incolore !
Exercice 5.3
:
On a obtenu, par mutagenèse d’une souche haploïde sauvage (SSR) de la
levure Saccharomyces cerevisiae, onze mutants indépendants, auxotrophes
pour le tryptophane.
1. Les onze mutants, croisés par la souche SSR, donnent des diploïdes
poussant sur milieu minimum avec ou sans tryptophane. Interprétez.
2. Les onze mutants sont croisés entre eux, sur un milieu additionné de
tryptophane, et donnent des diploïdes dont on teste le phénotype en les
répliquant sur milieu minimum Mo.
a. Justifiez ce protocole de croisement et de réplique.
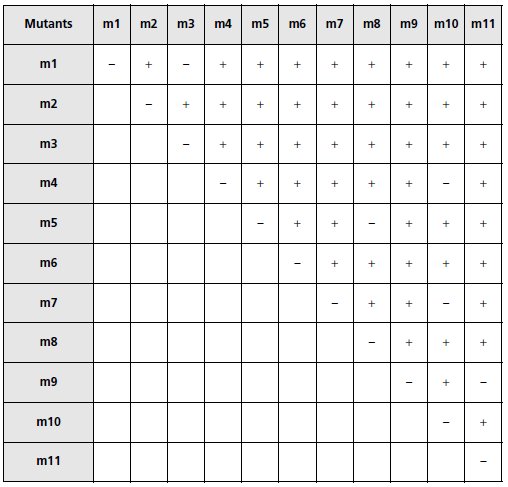
b. À partir des observations (tabl. 5.5) définissez les groupes de complémentation
réunissant les mutants ne complémentant pas entre eux.
TABLEAU 5.5 CROISEMENTS DE MUTANTS ENTRE EUX.
« + » signifie que le diploïde repiqué sur Mo est capable de croître et
« – » qu’il en est incapable.
c. Quelles interprétations simples peut-on formuler pour le mutant m10 ?
d. Quel est le moyen de vérifier laquelle correspond à la réalité ? (question
facultative.)
3. On dispose de boîtes de Pétri de milieu minimum avec très peu de tryptophane
(juste assez pour assurer une très faible croissance de mutants
auxotrophes, ne conduisant pas, normalement, à la formation d’un tapis cellulaire dense).
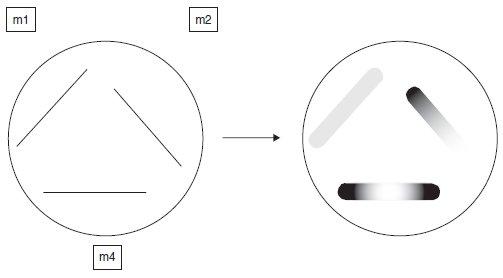
On ensemence une telle boîte avec des cellules de trois
souches m1, m2 et m4, et on obtient les résultats indiqués sur la figure 5.4,
sachant que les produits accumulés par le blocage d’une chaîne de biosynthèse
sont capables de diffuser dans le milieu, dans quel ordre les gènes interviennent-
ils dans la chaîne de biosynthèse ?
Figure 5.4 État de la boîte à l’étalement (à gauche);
après quelques jours (à droite).
– Maîtriser l’interprétation d’un tableau de résultat d’un test de complémentation
fonctionnelle.
– Définir la position d’un gène (de son produit) dans une chaîne.
Solution
1. Les diploïdes sont prototrophes, l’auxotrophie est récessive pour tous les mutants.
2. a. Justification du protocole : il s’agit, en croisant les mutants entre eux, de voir si le
diploïde est prototrophe pour le tryptophane, par complémentation fonctionnelle, ou auxotrophe,
par absence d’une telle complémentation, ce qui permettra de désigner les mutants
dans un même gène (groupes de complémentation) et d’estimer un nombre minimal de gènes
impliqués dans la biosynthèse de cet acide aminé.
Comme on ne sait pas à l’avance si le diploïde sera prototrophe ou auxotrophe, il convient de
le faire pousser d’abord sur une boîte de milieu additionné en trp, ce qu’il fera dans tous les
cas (proto ou auxo) puis de répliquer les colonies sur milieu minimum (sans trp) afin de voir
si elles poussent, et de tester ainsi leur prototrophie, c’est-à-dire la complémentation
fonctionnelle.
Si on avait directement étalé sur milieu minimum, la pousse de colonies aurait prouvé la
complémentation fonctionnelle, mais l’absence de colonies aurait pu s’interpréter aussi bien
comme une absence de complémentation fonctionnelle chez les diploïdes que comme une
absence de diploïdes tout court, par échec technique du croisement.
Remarque. Un résultat négatif est ambigu et difficile à interpréter.
Le protocole de
réplique utilisé permet de lever toute ambiguité, la présence de colonies sur le premier
milieu attestant de la réussite du croisement et de l’obtention de diploïdes.
b. On obtient 7 groupes de complémentation :
(m1, m3); (m2); (m4, m10); (m5, m8); (m6); (m7, m10); (m9, m11).
Il y a donc au moins sept gènes impliqués dans la chaîne de biosynthèse du tryptophane.
c. Il est important de noter que les mutants m4 et m7 complémentent entre eux et qu’ils ne
sont donc pas mutés dans un (le) même gène; en revanche, le mutant m10 apparaît dans deux
groupes de complémentation, ce qui signifie qu’il est obligatoirement muté dans au moins
deux gènes, au moins un gène en commun avec le mutant m4, et au moins un gène différent
du (des) précédent(s) en commun avec le mutant m7.
Il est possible de faire au moins deux hypothèses concernant le mutant m10, muté dans au
moins deux gènes :
– ce peut être un double mutant, les deux gènes étant touchés par deux mutations indépendantes;
– ce peut être un simple mutant, les deux gènes étant simultanément touchés par la même
mutation, ce qui est possible si les deux gènes sont contigus et que la mutation est une
délétion s’étendant sur les deux gènes en question.
d. On peut tester ces deux hypothèses par l’étude de la recombinaison génétique chez le
diploïde issu du croisement m10 × SSR :
– si m10 est muté dans deux gènes on doit observer, du fait de la recombinaison éventuelle,
des tétrades avec une seule spore sauvage (parentale) et trois spores de phénotype muté,
dont deux recombinées simple mutantes.
Dans le cas de l’analyse en vrac, on aura 3/4 de
spores mutées en cas d’indépendance mais la proportion diminuera d’autant plus vers 1/2
si les gènes sont liés;
– si m10 est un simple mutant par délétion dans deux gènes contigus, tout crossing-over
entre une chromatide sauvage et une chromatide délétée donnera une chromatide délétée et
une chromatide sauvage de sorte qu’on aura une stricte ségrégation 2/2; ce qui
pourrait encore être compatible avec l’hypothèse de deux gènes très proches.
3. Dans la mesure où il y a un peu de tryptophane, les cellules de chacun des trois sillons
commencent à pousser mais le tapis cellulaire stoppe son développement dès lors que le
milieu est épuisé, sauf en trois endroits, l’extrémité du sillon m2, du côté de m1, et les deux
extrémités du sillon m4.
Ce résultat signifie qu’une substance produite par m1 peut diffuser et permettre à m2 et à m4
de continuer à pousser et qu’une substance produite par m2 peut diffuser et permettre à m4
de pousser (l’inverse n’étant pas possible).
Sachant que les mutants auxotrophes ont une de leur étape de biosynthèse bloquée, et qu’ils
accumulent de ce fait l’intermédiaire produit en amont du point de blocage, on peut conclure
que m1 est bloqué plus en aval que m2 et m4, si bien que l’intermédiaire excrété par m1 et
fourni à m2 et m4 peut y être transformé en tryptophane tandis que les intermédiaires fournis
à m1 par m2 et m4 ne lui sont d’aucun secours. m2 est, lui, bloqué plus en aval que m4.
L’ordre des gènes dans la chaîne de biosynthèse est donc : m4-m2-m1.
Exercice 5.4
On a obtenu, par mutagenèse d’une souche haploïde sauvage (SSR) de la
levure Saccharomyces cerevisiae, quinze mutants indépendants, auxotrophes pour l’uracile. Un test de complémentation fonctionnelle a permis de
répartir ces mutants en 5 groupes de complémentation (1, 9, 11, 14, 15),
(2, 10), (3, 5, 6, 13), (4, 7, 15) et (8, 12).
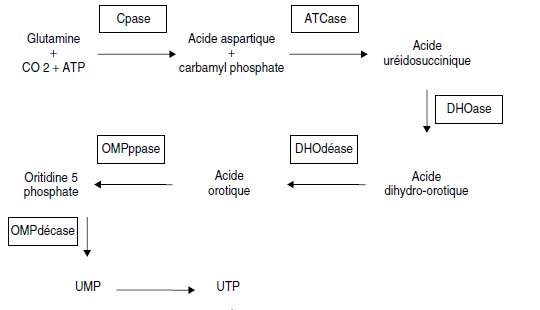
Par ailleurs, des études biochimiques ont permis de définir la chaîne de
biosynthèse de l’uracile, sous forme d’UTP, avec les principaux intermédiaires
et les activités enzymatiques associées aux différentes transformations,
selon la figure 5.5.
Figure 5.5.
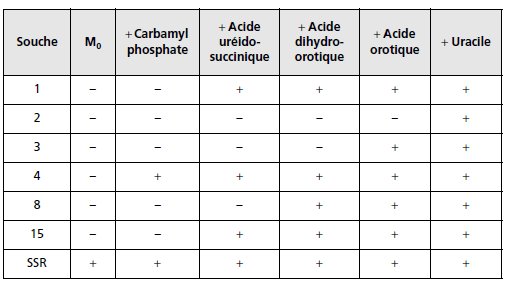
TABLEAU 5.6.
Capacité de croissance des souches en présence d’un intermédiaire.
« + » indique la présence et « – » l’absence de colonies.
1. Les souches mutantes 1, 2, 3, 4, 8 et 15 ont été mises à croître sur milieu
minimum ou milieu additionné d’un des métabolites intermédiaires de la
chaîne présentée ci-dessus. Vous interprétez, le plus complètement possible,
ces résultats (tabl. 5.6) en indiquant dans quel(s) gène(s) chaque mutant est
touché, particulièrement pour les mutants 1 et 15, compte tenu de la définition
des groupes de complémentation.
NB : en réalité ces différents produits intermédiaires, sauf l’uracile, ne
peuvent pénétrer dans la cellule.
On imaginera, pour le problème qu’ils le
peuvent…
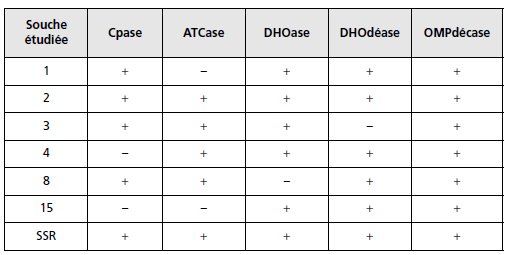
2. Il est possible de tester dans un extrait acellulaire plusieurs des activités enzymatiques
impliquées dans cette chaîne de biosynthèse (tabl. 5.7). Concluez.
TABLEAU 5.7 TEST DES ACTIVITÉS ENZYMATIQUES.
« + » indique une activité décelable.
Mettre en relation les résultats d’un test génétique de complémentation fonctionnelle
et ceux de tests biochimiques d’activité enzymatique ou trophiques de croissance
sur des milieux supplémentés.