1. Souche 1 : pousse avec acide uréidosuccinique, elle est donc mutée dans au moins un gène
gouvernant une étape antérieure, et dans aucun des gènes gouvernant les étapes postérieures.
Souche 2 : mutée dans une étape ultérieure, soit dans le gène de l’OMPppase, soit dans celui
de l’OMPdécase, cette étape n’ayant pas été testée isolément.
Souche 3 : mutée dans le gène de la DHOdéase.
Souche 4 : mutée dans le gène de la Cpase.
Souche 8 : mutée dans le gène de la DHOase.
Souche 15 : même réponse que pour la souche 1, soit pousse avec acide uréidosuccinique,
elle est donc mutée dans au moins un gène gouvernant une étape antérieure, et dans aucun
des gènes gouvernant les étapes postérieures.
La souche 15 est au moins un double mutant puisqu’elle est présente dans deux groupes
distincts de complémentation.
Ne complémentant pas avec la souche 4, on peut en déduire
que la souche 15 est mutée dans le gène de la Cpase, gène qui n’est pas touché dans la souche 1
qui complémente avec la souche 4.
Le gène qui est touché dans la souche 1 et dans la souche 15
est donc celui de l’ATCase; la souche 1 n’étant mutée que dans ce seul gène, sinon elle ne
complémenterait pas avec la souche 4.
Les souches 3, 4 et 8 sont des mutants simples, car s’ils étaient respectivement mutés dans
des gènes intervenant antérieurement à leur point de blocage, ils ne complémenteraient pas
entre eux.
2. Souche 1 : déficit en ATCase, confirmant les déductions précédentes (mutant simple).
Souche 2 : aucun déficit, mais l’enzyme OMPppase n’ayant pas été testée, on peut en déduire
que c’est un mutant simple dans le gène de cette enzyme (voir question précédente).
Souche 3 : déficit en DHOdéase, confirmant les déductions précédentes (mutant simple).
Souche 4 : déficit en Cpase, confirmant les déductions précédentes (mutant simple).
Souche 8 : déficit en DHOase, confirmant les déductions précédentes (mutant simple).
Souche 15 : double déficit confirmant le fait que le mutant 15 est un double mutant dans
les deux gènes de la Cpase et de l’ATCase (à moins que les deux activités enzymatiques
soient réalisées par la même chaîne peptidique, codée par un seul gène, et que les mutations
de ce gène gouvernant ces deux activités ne retentissent que sur l’une des deux activités
ou sur les deux activités selon le site de mutation dans le gène et l’effet de la
mutation !).
Exercice 5.5
On se propose d’étudier, chez la levure Saccharomyces cerevisiae, des
mutants de phénotype [gal–] incapables de métaboliser le galactose.
On dispose, pour cela, de deux souches haploïdes, A de signe sexuel a et
de phénotype [gal+; val–], auxotrophe pour la valine (val), et B de signe
sexuel α et de phénotype [gal+; trp–] auxotrophe pour le tryptophane (trp).
On obtient, par mutagenèse, des mutants [gal–], dont on vérifie qu’ils sont
restés [val–] ou [trp–].
On dispose ainsi de quatre mutants indépendants à partir de la souche A,
notés A1, A2, A3 et A4, et de quatre mutants indépendants à partir de la
souche B, notés B1, B2, B3 et B4.
On croise ces mutants entre eux ou avec une des deux souches A ou B,
puis on teste la capacité des diploïdes à pousser sur galactose.
1. Sur quel milieu obtenez-vous les diploïdes, avant de les tester pour le
phénotype gal ? Justifiez votre réponse en précisant le rôle des mutations
d’auxotrophie.
2. Comment testez-vous le phénotype gal ? Justifiez votre réponse.
3. Interprétez, avec des schémas clairs et démonstratifs, les résultats
obtenus (tabl. 5.8).
4. Que faudrait-il faire pour croiser entre elles des souches porteuses des
mutations affectant A2 et A3 ? Pourquoi ce croisement est-il utile à envisager
?
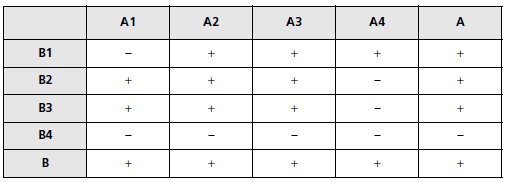
TABLEAU 5.8 PHÉNOTYPES DES DIPLOÏDES ISSUS DES CROISEMENTS ENTRE SOUCHES A ET B.
« + » désigne la capacité de croissance sur galactose.
– Maîtriser l’interprétation d’un tableau de résultat d’un test de complémentation
fonctionnelle.
– Comprendre le rôle des marqueurs de sélection des diploïdes et la contrainte du
signe sexuel nécessitant la construction de souches aptes à la réalisation des
croisements.
Solution
1. On peut croiser entre eux les mutants A et B, puisqu’ils sont de signe sexuel différent.
Comme ils sont [gal–], mais qu’on ne sait pas si ce phénotype mutant est dominant ou
récessif, il faut réaliser les croisements sur une boîte de milieu minimum avec glucose.
Le milieu utilisé est minimum, ce qui évite la pousse de colonies haploïdes qui sont [trp–]
ou [val–].
Seules des colonies de diploïdes peuvent se développer, puisque leurs cellules
peuvent croître et se multiplier par complémentation fonctionnelle pour ces mutations
d’auxotrophie qui jouent ici le rôle de marqueur de sélection des diploïdes.
2. On teste le phénotype gal par réplique sur une boîte de milieu minimum avec gal.
3. Le phénotype [gal–] du mutant B4 est dominant, les autres sont récessifs.
Les croisements
entre mutants récessifs réalisent un test de complémentation fonctionnelle et d’allélisme
pour les gènes impliqués dans le phénotype [gal–].
• Deux souches mutées dans un même gène donnent, par croisement, un diploïde où les
deux exemplaires de ce gène sont non fonctionnels, d’où un phénotype [gal–] (fig. 5.2 et
raisonnement associé).
• Deux souches mutées dans des gènes différents donnent, par croisement, un diploïde dont
le phénotype est [gal+], par complémentation fonctionnelle (fig. 5.1 et raisonnement associé).
L’application de ce principe aux résultats observés (tabl. 5.6) permet de définir 5 groupes de
complémentation, a : {A1; B1}, b : {A2}, c : {A3}, d : {A4, B2} et e :{A4, B3}.
Remarque 1. Un groupe de complémentation correspond, le plus souvent, à un gène
mais ce n’est pas obligatoire.
A3, par exemple, peut être un double mutant, dans ce
cas le groupe c correspondrait à deux gènes; le groupe a correspond plus vraisemblablement
à un gène car, dans le cas contraire, il faudrait supposer que A1 et B1,
mutants indépendants, soient simultanément mutés dans les mêmes gènes.
Remarque 2. Si A4, B2 et B3 sont des mutants simples (un seul gène) alors d et e ne
forment qu’un seul et même groupe de complémentation, sinon A4 est un mutant
double appartenant à deux groupes de complémentation différents.
Cette incertitude vient ici du fait que tous les croisements possibles entre mutants
n’ont pas été faits, notamment B2 × B3, puisque ceux-ci sont de même signe sexuel.
4. Il faudrait pouvoir disposer d’une souche porteuse de la mutation affectant B2 ou B3, mais
de signe sexuel a.
Ce croisement serait utile, car le TCF réalisé ici ne concerne pas tous les mutants obtenus
mais seulement ceux de signe sexuel opposé; il est donc impossible de dire si A2 et A3, ou
B2 et B3, appartiennent ou non au même groupe de complémentation.
Exercice 5.6
On dispose, chez la levure Saccharomyces cerevisiae, d’un mutant haploïde M,
incapable de métaboliser le galactose, phénotype noté [gal–], et d’une souche
haploïde sauvage S [gal+].
Question 1.
On effectue le croisement M × S et on vérifie que le diploïde obtenu est de
phénotype [gal+], puis on le met sur un milieu de sporulation qui permet
d’isoler 100 tétrades dont on fait l’analyse.
On observe :
– 75 tétrades de type 1 avec 2 spores [gal+] et deux spores [gal–];
– 20 tétrades de type 2 avec 1 spore [gal+] et trois spores [gal–];
– 5 tétrades de type 3 avec quatre spores [gal–].
Vous ferez une analyse génétique complète de ces résultats (avec génotypes).
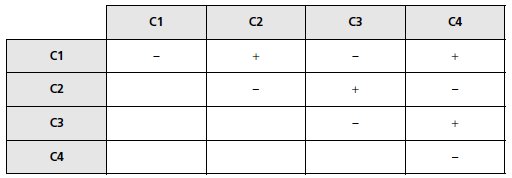
Question 2.
On récupère, dans une tétrade de type 3, les quatre cellules [gal–], désignées
par C1, C2, C3 et C4 et on s’arrange, après les avoir multipliées,
pour les croiser entre elles, ou avec des dérivées de même génotype sauf le
signe sexuel. On obtient les résultats suivants que vous interpréterez en
précisant le génotype de chacune d’entre elle (le signe « + » indique le
phénotype [gal+]).
NB : la situation des spores étant symétrique, vous ferez un choix arbitraire
pour leur génotype, mais vous conserverez évidemment ce choix
pour tout le reste du problème.
– Maîtriser l’interprétation fonctionnelle de la complémentation.
– Utiliser l’analyse de tétrades pour différencier des spores recombinées de même
phénotype mais de génotype différent.
Solution
1. Il n’y a pas de ségrégation 2-2 car dans ce cas, la méiose du diploïde donnerait toujours
deux spores [gal+] et deux spores [gal–] et on n’observerait qu’un seul type de tétrades. M
diffère donc de S pour au moins deux gènes, elle est donc de génotype (a; b)
Le croisement M × S s’écrit ainsi en phénotypes et en génotypes :
Souches croisées : M × S
Phénotypes parentaux : [gal–] × [gal+]
Génotypes parentaux : (a, b) × (A, B)
Génotype du diploïde : (A // a ---- B // b)
Génotypes des spores : (A, B) + (a, b) + (A, b) + (a, B)
* déductible de l’analyse des tétrades : il y a une tétrade avec quatre spores [gal–].
L’analyse de tétrades conduira à l’observation de DP avec quatre spores parentales, deux (A, B)
et deux (a, b), c’est-à-dire deux spores de phénotype [gal+] et deux spores de phénotypes
[gal–], et, si il y a recombinaison, des tétrades avec trois spores [gal+] et une spore [gal–],
enfin des DR avec quatre spores [gal–].
Par ailleurs comme on peut remarquer que la fréquence des DP est largement supérieure à
celle des DR, on peut conclure que les gènes A et B sont génétiquement, donc physiquement
liés, à une distance corrigée :
d = f(T)/2 + 3f(DR) = 25 ur
2. Les spores C1, C2, C3 et C4 sont toutes quatre recombinées et ne possèdent donc qu’une
seule mutation : elles sont (A, b) ou (b, A).
Le tableau est un test de complémentation fonctionnelle (on sait que le mutant M est récessif
et que les deux mutations a et b le sont donc aussi vis-à-vis de leurs homologues sauvages
respectifs) qui montre que C1 et C2 sont mutées dans des gènes différents, donc l’une C1 est
(a, B) alors que l’autre, C2 est (A, b).
Bien évidemment C1 et C3 ont même génotype et
symétriquement C2 et C4.
Cet exemple illustre l’un des intérêts de l’analyse de tétrades : en allant rechercher des spores
dans les DR, on sait qu’elles sont recombinées et simples mutantes, alors que leur phénotype
mutant n’aurait pas permis de les distinguer de spores parentales doubles mutantes, dans une
analyse de spores en vrac.
L’analyse de tétrades est un outil de tri à utiliser pour identifier les
spores recherchées dans l’exercice précédent.