Analyse fonctionnelle et moléculaire des révertants et des supresseurs :
A - Introduction :
Si le phénotype observé en aval d’une mutation est attaché à une perte de fonction
d’un gène, cette perte de fonction peut résulter de mutations très variées, à la fois
dans la séquence du gène affectée (promoteur, séquences 3′ ou 5′, intron, séquence
codante) que dans la manière dont elle est affectée (délétion, addition, substitution
de pb).
Parmi ces mutations, on peut citer les mutations ponctuelles de la séquence
codante, par substitution d’une paire de base par une autre (mutation non-sens ou
STOP et mutation faux sens) ou par décalage du cadre de lecture, et les mutations
d’épissage.
Cependant, il convient de distinguer, parmi les mutations de perte de fonction,
celles où le message génétique est modifié (mutations ponctuelles) et celle où il est
perdu (délétions) car cette distinction est critique dans la capacité d’avoir ou non un
suppresseur et dans la nature de ce suppresseur éventuel.
Dans le cas où la perte de fonction du gène résulte d’une modification ponctuelle
du message, on va voir qu’on peut obtenir des révertants vrais, ou des révertants par
une nouvelle modification ponctuelle du message qui rétablit une information fonctionnelle,
ou par un suppresseur dit informationnel car il a pour effet de corriger
l’erreur du message génétique dans le processus de son expression (épissage, traduction).
D’une manière ou d’une autre, la fonction du gène est rétablie chez le révertant.
Dans le cas où la perte de fonction du gène résulte d’une perte du message génétique
lui-même (délétion partielle ou totale de la séquence codante), la fonction du
gène ne peut être rétablie.
Il se peut alors qu’il n’y ait pas de révertant possible si
cette fonction est incontournable (c’est d’ailleurs un critère d’identification des mutants
par délétion) mais il se peut qu’on puisse sélectionner des révertants porteurs d’un
suppresseur.
Dans ce cas on va voir qu’on obtient un suppresseur dit fonctionnel ou
physiologique, car il ne rétablit pas la fonction perdue du gène mais corrige l’effet de
cette fonction perdue en la contournant, par exemple en la rendant inutile, ce qui
peut donc s’appliquer à toutes les mutations de perte de fonction de ce gène, qu’elles
soient ponctuelles ou que ce soit des délétions.
Dans le cas de mutations de gain de fonction, le message est modifié et conduit à
la présence d’une protéine mutée responsable du phénotype muté, les révertants seront
soit porteurs d’une nouvelle mutation du gène dont l’effet annule l’effet de la première,
soit d’une mutation dans un gène suppresseur dont le produit, par interaction avec le
produit muté, peut en annuler les effets phénotypiques.
B - Analyse et interprétation moléculaire des révertants
de première classe ou de certains révertants
de seconde classe avec un suppresseur très lié :
Quand il est possible d’étudier la chaîne peptidique produite chez des révertants de
première classe ou des révertants de seconde classe avec un suppresseur très lié, on
peut, en la comparant avec la chaîne peptidique sauvage obtenir l’un des quatre résultats
suivants :
– la chaîne peptidique des révertants diffère de la chaîne peptidique sauvage par un
et un seul acide aminé, variable d’un révertant à l’autre mais substituant toujours
le même acide aminé de la souche sauvage;
– la chaîne peptidique des révertants diffère de la chaîne peptidique sauvage par
plusieurs acides aminés contigus, en nombre variable d’un révertant à l’autre, en
amont ou en aval d’un même acide aminé;
– la chaîne peptidique des révertants diffère de la chaîne peptidique sauvage en
étant plus longue ou plus courte, du côté N-terminal, l’acide aminé en position 0
n’étant pas une méthionine;
– la chaîne peptidique des révertants diffère de la chaîne peptidique sauvage en
étant plus longue ou plus courte, du côté C-terminal.
Chacun de ces quatre cas admet une interprétation simple qui permet d’ailleurs de
préciser la position et la nature de la mutation directe affectant le mutant.
On remarquera, en conséquence, que selon la position et la nature de la mutation
directe, on n’obtient pas le même type de révertant de première classe ou de deuxième
classe avec suppresseur très lié.
a) Les révertants dont la chaîne peptidique diffère
de la chaîne sauvage par un et un seul acide aminé :
Il s’agit de révertants où une deuxième mutation survient au site de la mutation
directe.
Ces révertants sont issus de mutants directs ponctuels, chez lesquels une
paire de base avait été substituée par une autre.
Cette substitution peut avoir transformé
un codon sens en un codon stop (mutations non sens), ce qui se traduit par une
perte de fonction du gène, son produit étant absent par arrêt de la lecture.
Cette substitution
peut aussi avoir changé le sens du codon (mutation faux-sens).
Le phénotype
muté, chez le mutant direct, peut être dû à une perte de fonction (produit inactif), ou
à des effets plus subtils (produit plus actif ou moins actif ou doué d’une autre activité
biologique).
Un révertant au site muté a subi, dans le codon muté, une nouvelle substitution
d’une paire de bases par une autre :
– à partir d’un mutant direct de type stop (non-sens), un révertant au site muté présentera
un nouveau codon signifiant, différent de sauvage, ce qui conduit à la présence
d’un acide aminé différent de celui de la séquence peptidique sauvage, mais compatible
avec la fonction de la protéine, puisque le révertant est de phénotype sauvage.
Exemple :
– chez la SSR, en un site du gène, il y a un codon TGG (trp),
– chez le mutant direct, le codon muté est TAG (stop),
– chez le révertant vrai, le codon TAG est redevenu TGG,
– chez un révertant muté au site, le codon TAG peut être muté en GAG (glu), ou AAG (lys), ou…,
– à partir d’un mutant direct de type faux-sens, un révertant au site muté présentera
un nouveau codon signifiant, différent du codon sauvage, ce qui conduit à la
présence d’un acide aminé différent de celui de la séquence peptidique sauvage
mais aussi de la séquence peptidique mutée, et, cette fois, compatible avec la fonction
de la protéine, puisque le phénotype du révertant est sauvage.
Exemple :
– chez la SSR, on a un codon TGG (trp), spécifiant un acide aminé très hydrophobe,
– chez le mutant direct, le codon est muté CGG (arg), spécifiant un acide aminé
basique incompatible avec la structure tri-dimensionnelle de la protéine,
– chez le révertant vrai, le codon CGG est redevenu TGG,
– chez un révertant muté au site, le codon CGG est muté en GGG (gly) compatible
avec la structure.
En conclusion les révertants de première classe peuvent être identifiés comme des
révertants au site muté par l’analyse comparative des chaînes peptidiques sauvages
et révertantes.
Remarque 1. On peut très éventuellement imaginer, chez le diploïde issu du
croisement révertant par sauvage, une recombinaison intra-codon reconstituant
un gène muté, si la mutation directe touche l’une des paires de base alors que
la mutation réverse, redonnant un codon sens ou un codon compatible, touche
une autre paire de base du codon que la mutation directe (voir exercices et
remarque 4, plus haut).
Remarque 2. La sélection de révertants au site (codon) de la mutation directe
et l’analyse des peptides sauvage et révertant ont joué un rôle historique
important dans la compréhension de la relation gène-peptide et le codage de
l’information génétique.
À partir d’un grand nombre de mutants ponctuels du
gène de la tryptophane synthétase de coli et des révertants au site muté pour
chacun de ces mutants, on a pu établir la « colinéarité gène-protéine », à savoir
que la position relative des mutations dans le gène correspondait à la position des acides aminés modifiés
dans la chaîne peptidique.
Ce résultat a conforté l’idée que l’ordre d’enchaînement
des acides aminés était spécifié par l’ordre d’enchaînement des
nucléotides puis au concept de codon puisqu’il n’y avait que quatre types de
bases pour vingt acides aminés différents.
b) Les révertants dont la chaîne peptidique diffère de la chaîne sauvage
par plusieurs acides aminés contigus :
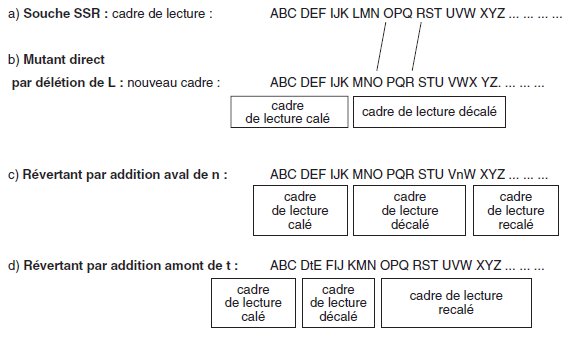
Il s’agit en général de révertants issus de mutants directs par décalage du cadre de
lecture. Une deuxième mutation est survenue chez le révertant, non loin de la
première et de signe opposé (une addition si la mutation directe est une délétion,
une délétion si la mutation directe est une addition) qui a pour effet de recaler le
cadre.
La séquence nucléotidique est dans le cadre normal avant la première mutation et
après la seconde, et reste décalée entre les deux sites mutés, ce qui conduit à un
changement de la séquence peptidique entre ces deux points.
Évidemment les deux mutations, directe de décalage, et suppresseur de recalage,
ne peuvent être trop éloignées l’une de l’autre sinon la séquence peptidique aberrante
serait trop longue et incompatible avec une conformation fonctionnelle de la
protéine.
La mutation suppresseur peut survenir en aval ou en amont de la mutation directe;
dans le premier cas la mutation suppresseur recale le cadre; dans le deuxième cas la
mutation directe qui décalait le cadre, en absence de la mutation suppresseur, le
recale désormais (figure 7.2).
La comparaison des séquences peptidiques entre la chaîne sauvage et la chaîne révertante, permet de préciser la position et la nature de la mutation directe, éventuellement
celle de la mutation suppresseur, ainsi que la séquence nucléotidique du
gène entre les deux sites mutés, malgré la dégénérescence du code (voir exercices).
Figure 7.2 Mutation directe de décalage du cadre de lecture
et suppresseur intragénique de recalage en aval
ou en amont de la mutation directe.
c) Les révertants dont la chaîne peptidique diffère de la chaîne sauvage
par leur séquence N-terminale ou C-terminale :
Il s’agit de révertants issus de mutants directs au site d’initiation ou au site de terminaison
de la lecture.
La mutation du site AUG d’initiation de la lecture peut aboutir
à l’absence de chaîne peptidique si il n’y a pas d’autre site AUG en phase (dans le
même cadre de lecture) au voisinage du site naturel, ou à une chaîne « mutante »
(non fonctionnelle ou modifiée) si un tel site existe.
Le révertant est alors un mutant qui présente un nouveau site AUG, en phase, dans
le voisinage du site naturel, de telle sorte que la chaîne peptidique est fonctionnelle,
ce qui conduit à un phénotype sauvage.
Si le site est en 3′ du site naturel, la chaîne révertante est plus courte que la chaîne sauvage de quelques acides aminés, si le site
est en 5′ du site naturel, la chaîne révertante est plus longue de quelques acides
aminés, ce qui permet alors de spécifier, par le code génétique de l’acide aminé en
position 0, la nature de la mutation directe affectant le codon AUG (voir exercices).
Le même type d’analyses a été développé pour les mutations directes affectant le
codon stop dans les gènes α ou β de l’hémoglobine
d) Les cas particuliers
:
Ils sont nombreux et on en citera trois.
➤ Double mutation faux-sens dans la séquence codante
On peut donner l’exemple de la tryptophane synthéthase de coli.
Le révertant sauvage
présente une activité enzymatique, absente chez le mutant.
L’analyse de la chaîne
peptidique révertante montre la substitution de deux acides aminés de la séquence
sauvage (glu et cys) par deux autres acides aminés (gly et tyr), les deux sites de
mutation faux-sens étant assez éloignés.
Ce résultat amène à conclure que le couple d’acides aminés glu-cys doit jouer un
rôle dans la conformation tridimensionnelle et/ou l’activité de la protéine et que la
mutation faux-sens directe de glu en gly ou de cys en tyr entraîne une instabilité et/ou
une perte de fonction.
L’effet suppresseur d’une deuxième mutation faux sens provient du fait que le
couple gly-tyr a un effet suffisamment semblable à celui du couple glu-cys pour rétablir
la stabilité et/ou l’activité protéique.
On a effectivement vérifié que les gènes porteurs de l’une ou de l’autre des deux
mutations faux-sens étaient non fonctionnels.
➤ Mutations directes dans le promoteur
La perte de fonction d’un gène peut résulter d’une mutation dans une séquence autre
que sa séquence codante, par exemple dans le promoteur.
Dans un tel cas les révertants
de première classe, qui expriment de nouveau le gène, sont mutés dans le
promoteur de telle sorte que la transcription est de nouveau possible.
Bien évidemment la séquence peptidique chez tous ces révertants est toujours
identique à celle de la SSR, ce qui distingue les mutants dans le promoteur des mutants dans la séquence codante pour lesquels seuls les révertants vrais présentent une
séquence peptidique sauvage.
➤ Fusion de promoteur
Le message du gène peut être inaccessible (ou trop accessible) si une mutation
affecte le promoteur du gène.
Dans ce cas la fonction du gène est altérée d’un point
de vue strictement quantitatif; la fonction est perdue si le gène ne peut plus être
suffisamment transcrit.
La restauration d’un phénotype normal dépendra soit d’une
nouvelle mutation du promoteur, rendant le message codé de nouveau accessible à
l’expression, par la transcription, soit d’un suppresseur fonctionnel, s’il peut en
exister un.
Dans ce cas un troisième type de révertant peut être sélectionné, si une délétion
fusionne la séquence codante du gène muté avec le promoteur d’un gène en amont
(dont la séquence codante de ce gène se trouve de fait déletée).
Dans tous les cas, les révertants présenteront une chaîne peptidique sauvage puisque
la séquence codante du gène n’était pas modifiée chez le mutant mais simplement
inaccessible.
C - Les suppresseurs informationnels
:
Un suppresseur informationnel rétablit la fonction du gène muté en agissant dans
son processus d’expression de manière telle qu’il y a production d’un produit fonctionnel
du gène, alors que celui ci demeure toujours muté.
Ainsi, d’une manière ou
d’une autre le processus d’expression, transcription, épissage, traduction, va à son
terme avec au bout une chaîne peptidique fonctionnelle, en tout cas assez pour
redonner un phénotype sauvage.
a) Les ARN-t suppresseurs informationnels
:
Il est assez facile d’obtenir chez les micro-organismes, bactéries ou levure, des
ARN-t suppresseurs informationnels, chez les révertants issus de mutants directs de
type stop (UAG, UAA ou UGA).
Ces ARN-t suppresseurs sont pourvus, par mutation
de leur séquence codante, d’un anti-codon complémentaire du codon stop correspondant
à la mutation directe.
De ce fait l’ARN-t muté-suppresseur devient capable de se positionner sur ce codon
stop, d’y apporter un acide aminé, ce qui permet d’éviter l’arrêt de traduction qui
survenait chez le mutant, puis de la continuer jusqu’à la fin du messager.
Exemples :
– le gène sauvage contient un codon TAC (tyr), UAC sur l’ARN-m, reconnu par un
ARN-t portant l’anti-codon 5′GUA3′. [N.B. : Hybridation anti-parallèle]
– le gène muté est porteur d’un codon TAA, à la place de TAC, donnant UAA sur
l’ARN-m, et induisant un arrêt de traduction.
– le révertant est muté dans un gène d’ARN-t de tyrosine, dont l’anti-codon 5′GUA3′
est muté en 5′UUA3′, ce qui lui permet de s’hybrider au codon UAA et d’apporter
une tyrosine, en restaurant une séquence peptidique sauvage.
– un autre révertant est muté dans un gène d’ARN-t de glutamique, dont l’anti-codon
5′UUC3′ (reconnaissant le codon GAA) est muté en 5′UUA3′, ce qui lui permet
de s’hybrider au codon UAA et d’apporter un glutamique, en restaurant une séquence
peptidique complète, mais modifiée en un point par la substitution d’une tyrosine
par un glutamique, sans doute compatible avec la structure et l’activité de la chaîne
peptidique, puisque le révertant a été isolé.
La fonction du gène, perdue chez le mutant, est ainsi restaurée, à condition que
l’acide aminé apporté par l’ARN-t suppresseur, qui n’est pas forcément l’acide aminé
sauvage, soit compatible avec une conformation tridimensionnelle active de la protéine;
tous les suppresseurs potentiels sur le plan traductionnel ne sont pas forcément des
suppresseurs efficaces sur le plan fonctionnel.
Remarque 1. On comprend qu’un ARN-t suppresseur soit allèle-spécifique, il
est incapable de corriger d’autres mutations que le codon stop dont il est
spécifique; par contre il est capable de corriger l’effet de cette mutation stop
dans n’importe quel autre gène que le gène porteur de la mutation directe (voir
exemple val1-1 et ade2-1, plus haut).
Remarque 2. Le fait qu’un gène d’ARN-t mute pour donner un ARN-t
suppresseur est sans conséquence pour la traduction dans la mesure où les
gènes d’ARN-t sont très redondants (plusieurs dizaines ou centaines de copies
par génome).
Si un ARN-t porteur de l’anti-codon 5′UUC3′ (reconnaissant le codon GAA,
glu) est muté en 5′UUA3′, ce qui lui permet de s’hybrider au codon UAA et
d’y apporter un glutamique, les codons GAA des messagers continueront
d’être traduits par tous les ARN-t codés par les autres copies non mutées du
gène de cet ARN-t.
Remarque 3. Une partie des gènes a un triplet de terminaison de type TAA, UAA
sur le messager.
La présence d’un ARN-t suppresseur capable de s’hybrider à
un tel triplet peut conduire à un prolongemesnt anormal de la traduction de
ces messagers et à la formation de chaînes peptidiques alongées dans leur
pertie C-terminale et éventuellement mal conformée et/ou inactive ou toxique.
La présence d’un ARN-t suppresseur informationnel est donc potentiellement
toxique pour la cellule, à moins que la concentration de cet ARN-t suppresseur
soit assez faible pour que son effet toxique sur les codons stop UAA des
gènes non mutés soit négligeable, mais quand même assez élevée pour que
son effet suppresseur sur la mutation directe UAA du gène muté soit assez
efficace.
b) Le suppresseur informationnel du phénotype [sable] chez Drosophila
:
Le mutant [sable] apparu chez Drosophila a un corps jaune clair qui le distingue du
phénotype gris sauvage de référence.
Son analyse génétique par croisement avec la SSR montre que le mutant est récessif et muté dans un seul gène, autosomique.
On a obtenu un révertant qui croisé avec la souche SSR donne en F2 issue d’un
croisement F1 × F1, 1/4 de [sable] chez les mâles et 1/8 chez les femelles.
Le retour
du phénotype mutant prouve l’existence d’un suppresseur et les proportions observées
montrent qu’il est dominant et localisé sur le chromosome X (refaire les croisements
à titre d’exercice).
L’analyse moléculaire, chez le révertant, du produit du gène, absent chez le mutant,
permet de conclure à la présence d’une protéine sauvage; l’analyse moléculaire du
gène muté a permis de montrer qu’il était inactivé par insertion d’une séquence de
type transposon et que la mutation à effet suppresseur affectait un gène spécifiant une
protéine impliquée dans l’épissage, de manière telle que le transposon est « reconnu »
comme un intron et excisé comme tel.
Ce suppresseur est bien informationnel
puisqu’il permet de reconstituer, dans le processus d’expression du gène, une séquence
fonctionnelle.