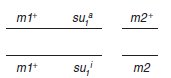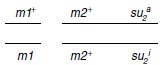Un suppresseur fonctionnel ou physiologique n’existe que si la physiologie cellulaire
peut être modifiée de façon telle que par son effet :
– le produit du gène porteur de la mutation directe devient facultatif ou inutile, dans
le cas où la mutation directe est une perte de fonction;
– le produit muté du gène voit son action contrecarrée par le produit muté du gène
suppresseur, dans le cas où la mutation directe est un gain de fonction.
Si le produit du gène est absolument nécessaire à la cellule, il n’y a pas de suppresseur
physiologique possible et des mutants par délétion ne présenteront jamais de
révertants.
En partant de ce principe, on peut considérer qu’un mutant ne donnant
pas de révertant est sans doute un mutant par délétion, puisque, pour tout autre type
de mutation, on peut potentiellement isoler des révertants.
a) Les suppresseurs fonctionnels de mutations de perte de fonction
:
Chaque cas est évidemment particulier puisqu’il tient à la fonction du gène affecté
par la mutation directe.
On peut cependant décrire des mécanismes observés à plusieurs
reprises.
– Mutation de surexpression d’un gène redondant ou homologue peu transcrit dans
le génotype sauvage.
Un certain nombre de fonctions biologiques peuvent dépendre
de deux ou plus de deux gènes dont les produits sont identiques (gènes redondants,
par exemple gènes α ou γ de l’hémoglobine) ou fortement homologues (par
exemple gènes des isozymes) de sorte que l’effet d’inactivation de l’un d’eux par
la mutation directe de perte de fonction, peut être supprimé par une mutation de
gain de fonction affectant un gène redondant ou homologue (voir les exercices sur
iso1 et iso2 cytochrome-c).
– Mutation activant une chaîne métabolique secondaire pouvant alors suppléer au
blocage de la chaîne principale du fait de la mutation directe.
Ce phénomène, sans être systématique est assez courant dans les réseaux métaboliques qui font un peu
penser à celui du chemin de fer : si une grande ligne est bloquée, avec un peu
d’astuce, on peut faire passer les trains par les voies secondaires.
– Court-circuit d’une cascade génique par mutation constitutive d’un gène s’exprimant
en aval du gène affecté par la mutation directe.
La levure diploïde est obligatoirement
hétérozygote pour le signe sexuel, ce qui conduit à la formation d’un hétérodimère par association des deux chaînes codées par les séquences Matα et
Mata.
Cet hétérodimère active une cascade d’expression de gènes conduisant à
l’activation du gène IME1 dont l’expression induit la méiose.
On a obtenu des
mutants de sporulation, mutés de façon telle que l’allèle muté Matam permet la
fécondation avec une souche Matα mais que le diploïde résultant est incapable de
sporuler car incapable d’induire une méiose.
L’obtention de révertants capables
de sporuler a montré, par analyse génétique, qu’ils étaient porteurs d’une mutation
suppresseur par gain de fonction rendant constitutif le gène IME1, ce qui rend
inutile la fonction du dimère.
– Mutation d’un gène régulateur au sein d’un réseau de régulation rendant inutile la
fonction du gène affecté par la mutation directe et permettant la reprise de
l’expression des gènes de structures.
Le mutant de drosophile NANOS est incapable
d’assurer la différenciation des segments abdominaux terminaux.
L’analyse
de ce mutant a montré qu’il était muté dans un seul gène.
On a obtenu un révertant
capable d’assurer un développement complet et le croisement du révertant avec
une souche sauvage a permis de retrouver en F2 des mutants, attestant de la
survenue d’un suppresseur.
L’analyse moléculaire du mutant et du révertant a
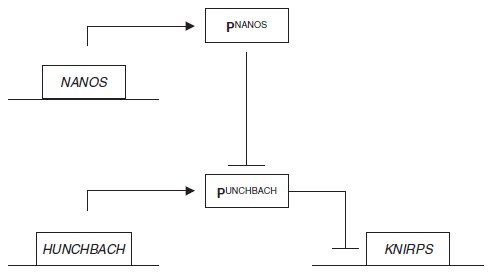
permis d’identifier les gènes impliqués et leurs fonctions respectives (figure 7.3) :
le gène NANOS code pour le répresseur du produit du gène HUNCHBACH qui,
lui même code pour le répresseur du gène KNIRPS qui active la différenciation
des segments abdominaux terminaux.
Le mutant NANOS étant un mutant de
perte de fonction dans ce gène, le produit du gène HUNCHBACH peut de
manière permanente réprimer le gène KNIRPS induisant alors l’incapacité de
différenciation des segments abdominaux terminaux.
Le révertant est un mutant
de perte de fonction dans le gène HUNCHBACH qui a pour conséquence de
rendre sans effet la perte de fonction dans le gène NANOS puisqu’il n’y a plus
de produit HUNCHBACH à réprimer, le gène KNIRPS pouvant alors s’exprimer
de manière constitutive (non régulée) sans que cela ait des effets néfastes sur le
développement embryonnaire.
On remarquera, au passage, que le révertant a
permis d’identifier le gène HUNCHBACH qui n’aurait jamais pu être identifié
par une mutation directe de perte de fonction qui eut été sans conséquence
phénotypique.
En effet la conséquence phénotypique de la perte de fonction de HUNCHBACH n’est phénotypiquement perceptible que dans le contexte de la
perte de fonction du gène NANOS.
Remarque 1. La sélection de révertants avec un suppresseur physiologique est
un bon moyen de toucher et d’identifier des gènes de régulation d’un gène de
structure.
Figure 7.3
b) Les suppresseurs fonctionnels de mutations de gain de fonction
:
Chaque cas est évidemment particulier puisqu’il tient à la fonction du gène affecté
par la mutation directe.
On peut cependant décrire des mécanismes observés à plusieurs
reprises.
➤ Mutation de perte de fonction dans le gène d’un répresseur muté
par une mutation directe de gain de fonction
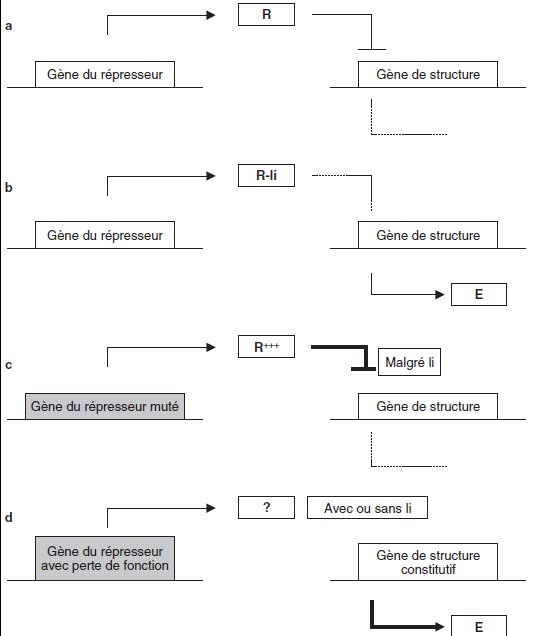
Exemple : l’expression d’un gène de structure, codant une enzyme E, est régulée par
un répresseur R, codé par le gène de celui-ci.
Le répresseur R bloque la transcription
du gène de structure (figure 7.4/a), à moins que, en présence d’un ligand li, R ne se
fixe plus sur le promoteur du gène de structure, ce qui permet alors son expression et
la présence de E (figure 7.4/b).
Un mutant, dépourvu de l’enzyme E, est muté, non dans le gène de structure, mais
dans le gène de son répresseur, de façon telle que le site de fixation n’a plus d’affinité
pour le ligand, le répresseur muté R+++ est « indétournable » de sa cible sur le
promoteur de enz, qui est définitivement bloqué (figure 7.4/c).
On isole un révertant où le gène régulateur est muté ponctuellement ou délété, le
répresseur n’existe plus et le gène enz devient constitutif, il est exprimé en présence
et même en absence du ligand li, mais il n’est plus régulé (figure 7.4/d).
➤ Expression anormale d’un partenaire du produit du gène porteur
de la mutation directe
De nombreux phénomènes comme la régulation du cycle cellulaire, les processus de
développement, l’homéostase, l’apoptose, la carcinogenèse, le vieillissement résultent
de l’expression et de l’action concertée de partenaires protéiques capables d’interagir.
La recherche de révertants peut être mise à profit pour mettre en évidence ce type d’interaction
et identifier les partenaires. Le gène BAX est un facteur pro-apoptotique retrouvé aussi bien chez l’homme que chez Drosophila ou Caenorhabditis elegans.
Pour rechercher des partenaires au produit du gène BAX, l’homme n’est certainement
pas un organisme modèle, ni Caenorhabditis elegans si on considère que de
nouveaux partenaires sont apparus dans l’évolution.
Figure 7.4
C’est pourquoi des chercheurs
ont imaginé d’utiliser cette bonne vieille drosophile comme outil d’investigation.
Ils
ont construit une souche transgénique :
– porteuse d’une insertion /pvg::GAL4/ (pvg est le promoteur du gène impliqué
dans le phénotype vestigial, de sorte que le produit PGAL4 du gène GAL4 ne s’exprime que dans les tissus de l’aile; PGAL4 est l’activateur de plusieurs des gènes
de levures impliqués dans la métabolisation du galactose);
– porteuse également d’une insertion /UAS::BAX/ (UAS étant la séquence cible de la
protéine PGAL4 au niveau des gènes de levure régulée par GAL4, de sorte que BAX
est exclusivement exprimé dans le tissu alaire).
Cette souche ainsi construite
présente un phénotype d’ailes avortées du fait de l’expression du facteur BAX proapoptotique.
L’avantage de cette construction est de permettre de confiner l’expression du gène
dans un tissu sans toucher à la viabilité de l’organisme qui reste disponible pour
transmettre les mutations et faire les croisements que nécessite l’analyse génétique.
Pour rechercher des partenaires de BAX, le mutant construit va être soumis à une
mutagenèse par insertion aléatoire d’éléments transposables P de drosophile porteurs
d’une séquence UAS. L’élément P va, un peu comme un rétrovirus, activer, par sa
séquence UAS (soumise à l’action de PGAL4) un ou des gènes dans le voisinage de
son site d’insertion.
Le but est alors de rechercher des révertants qui, étant pourvus
d’ailes plus ou moins normales seraient indicateurs de l’expression d’un partenaire
de BAX réprimant sa fonction pro-apoptotique.
Conclusions
:
L’analyse des révertants issus d’un ensemble mutants de même phénotype ouvre de
nombreuses portes.
– C’est d’abord un moyen d’étude de la mutation directe quand l’analyse de la chaîne
peptidique révertante est possible.
– C’est aussi un moyen de repérer les mutants par délétion quand aucun révertant,
notamment physiologique, ne peut être sélectionné.
– C’est un moyen de toucher de nouveaux gènes impliqués dans le phénomène
étudié et qui n’avaient pas été ou ne pouvaient être sélectionnés par le crible de
mutants directs, notamment certains gènes de régulation dont les mutations ne
peuvent être phénotypiquement perçues dans un contexte sauvage, et d’ouvrir la
voie à une étude du fonctionnement intégré des gènes.
– C’est enfin le moyen de définir un crible de mutants, soit pour rechercher des
partenaires au produit d’un gène donné, soit pour obtenir des mutants de perte de
fonction dans des gènes essentiels de l’activité cellulaire dont les mutations sont
létales et pour lesquels il ne semble pas possible d’obtenir des mutants de type
conditionnels (voir exercice 8.5).
La sélection et l’analyse des révertants s’avèrent donc être un outil puissant de
l’analyse génétique dans tous ses aspects, le criblage des mutants et l’identification
des gènes, leur analyse fonctionnelle et leur cartographie.
Evercices
:
Exercice 7.1
On ne tient compte dans ce problème, ni du signe sexuel des souches ni des
marqueurs de sélection des diploïdes.
On dispose, chez Saccharomyces cerevisiae, de deux mutants m1 et m2,
auxotrophes pour la valine, phénotype noté [val–].
Des analyses antérieures
ont montré que les deux phénotypes [val–] sont récessifs, que
chaque mutant m1 ou m2 n’est muté que dans un seul gène et que les deux
gènes mutés chez m1 et m2 sont physiquement indépendants.
1. On dépose environ 108 cellules du mutant m1 sur une boîte de milieu
minimum, après culture en milieu liquide, centrifugation et récupération
des cellules; on observe quelques clones, dont l’un est nommé c1.
On
renouvelle le même protocole à partir d’une deuxième culture, et on isole
un deuxième clone, nommé c2.
Interpréter ce résultat.
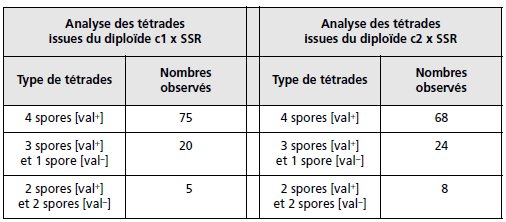
2. On croise chacun des clones c1 et c2 avec la SSR, les diploïdes sont mis
à sporuler et on analyse cent tétrades.
Interprétez l’ensemble de ces résultats
(tabl. 7.4) en justifiant vos réponses.
TABLEAU 7.4 CROISEMENT DES CLONES AVEC LA SSR.
3. On récupère les spores de phénotypes [val+] dans les asques contenant
2 spores [val+] et 2 spores [val–] à l’issue des croisements précédents
(tabl. 7.4).
Ces spores sont nommées v1 ou v2 selon qu’elles proviennent
de tétrades issues des croisements c1 × SSR ou c2 × SSR.
a. Quel est le génotype des spores v1 ou v2 ? Justifiez (éventuellement
après la question b) le fait d’avoir sélectionné ces spores [val+] dans ces
asques et non dans d’autres.
b. Les spores v1 sont croisées avec le mutant m2, et les spores v2 sont croisées
avec le mutant m1.
Les diploïdes sont mis à sporuler et on réalise l’analyse des tétrades issues de la méiose.
Vous interpréterez ces résultats
(tabl. 7.5) sur le plan cartographique et sur le plan fonctionnel (il est utile
de se souvenir de la cartographie établie), en essayant de dégager toutes les
conclusions possibles sur la nature des mutations en causes, et les mécanismes
moléculaires sous-jacents aux phénotypes observés dans les
tétrades obtenues.
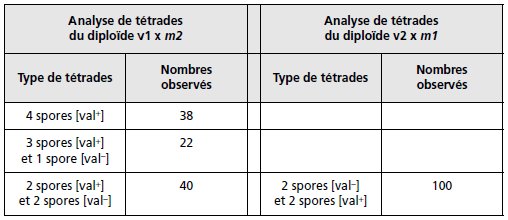
TABLEAU 7.5 CROISEMENT DES SPORES AVEC LES MUTANTS.
– Analyse de révertants, mise en évidence et cartographie des suppresseurs.
– Analyse fonctionnelle des mutations directes et des suppresseurs.
Solution
1. c1 est un révertant [val+], soit un révertant vrai, ce qui est rare, soit plutôt un révertant avec
une deuxième mutation à effet suppresseur (intragénique ou extragénique et, dans ce cas,
informationnelle ou physiologique).
Même réponse pour c2.
c1 et c2 sont des révertants indépendants et leur site de mutation, comme le mode d’action du
suppresseur (s’il y en a un) peuvent être différents.
Remarque. m1 et m2 sont porteurs de mutations ayant la même conséquence fonctionnelle,
l’auxotrophie pour la valine, mais les mutations affectant chacun des deux
gènes peuvent être de nature tout à fait différente (NS; FS; décalage; délétion, etc.) de
sorte que l’éventuel suppresseur de c1 qui est actif sur m1 peut être, ou ne pas être,
actif sur m2.
De la même façon, l’éventuel suppresseur de c2 qui est actif sur m2 peut
être, ou ne pas être, actif sur m1; telles sont les questions qu’on doit avoir à l’esprit si
on anticipe le déroulement de l’analyse.
2. Le croisement d’un révertant par la SSR est destiné à vérifier l’existence postulée d’un
suppresseur.
En effet, si un suppresseur existe et qu’il est suffisamment distant de la mutation
d’auxotrophie originelle, la méiose du diploïde révertant × SSR doit laisser réapparaître des
génotypes et donc des phénotypes mutants.
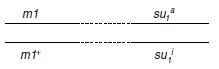
Le génotype du diploïde peut s’écrire, pour les gènes qui nous intéressent :
et on voit bien que la recombinaison, par assortiment aléatoire des chromosomes, si m et su1
sont physiquement indépendants, ou par crossing-over, si m et su1 sont physiquement liés,
doit donner (sauf s’ils sont très liés) des spores (m1, su1i) de phénotype [val–].
On attend, pour les DP, quatre spores [val+]; pour les T, trois spores [val+] et une spore [val–];
pour les DR, deux spores [val+] et deux spores [val–].
En effet, si les génotypes (m1+; su1i)
avaient un autre phénotype que [val+], le résultat observé serait tout autre (voir page 176).
L’analyse de tétrades montre donc, avec l’apparition de spores [val–] à l’issue des deux croisements,
que c1 et c2 sont des révertants porteurs d’une mutation à effet suppresseur, que le
suppresseur su1 est génétiquement lié au gène muté chez m1, et que le suppresseur su2 est
génétiquement lié au gène muté chez m2, puisque, dans chaque croisement, f(DP) > f(DR).
Comme m1 et m2 sont physiquement indépendants, on peut en déduire que les sites su1
et m1 (souche c1) sont physiquement indépendants des sites su2 et m2 (souche c2).
On peut,
par ailleurs, calculer les distances respectives des sites m1 et su1, d’une part (25 ur) et des
sites m2 et su2, d’autre part (36 ur) en appliquant la formule d = 100[3f(DR) + f(T)/2].
3. a En allant chercher des spores [val+] chez des DR, on peut définir sans ambiguité leur
génotype.
En effet, elles sont obligatoirement porteuses du suppresseur actif et du gène non
muté, ce qui est recherché pour la suite de l’analyse génétique.
On a donc :
– spore v1 génotype (m1+; su1a), qui est par ailleurs (m2+);
– spore v2 génotype (m2+; su2a), qui est par ailleurs (m1+).
Les tétratypes n’étaient pas utiles ici car les spores [val+] qu’ils renferment ont deux génotypes
possibles, (m+, sua) et (m+, sui), ce qui ne permet pas de faire les croisements suivants
avant d’avoir identifié laquelle des deux spores a le génotype recherché (m+, sua). Cette
opération aurait été nécessaire si on avait récupéré les spores en vrac.
Ici l’analyse de tétrades
nous en dispense.
3. b Les croisements réalisés donnent les diploïdes suivants :
– diploïdes v1 × m2
– diploïdes v2 × m1
Le but du jeu est à l’évidence dans le premier croisement, de réunir, par recombinaison à
l’issue de la méiose, la mutation m2 avec le suppresseur su1a de m1, afin de savoir s’il est
aussi actif sur m2.
Si c’est le cas, les spores de génotype (m2; su1a) seront de phénotype [val+], dans le cas
contraire, elles seront [val–]; et on sait que les spores de génotype (m2; su1a) seront réalisées,
notamment dans les nombreux DR qui doivent se former, puisque m2 et su1 sont physiquement
indépendants.
Comme on observe 38 tétrades avec quatre spores [val+], on peut en conclure que les deux
spores contenant m2 (ségrégation 2/2) sont [val+] parce qu’elles contiennent su1a qui est
donc actif sur m2.
On n’observe jamais, à l’issue des méioses du diploïde v2 × m1, de
tétrades avec quatre spores [val+].
Or on sait, du fait de l’indépendance physique entre les
sites m1 et su2a que les DR sont fréquents et que de nombreuses spores (m1; su2a) sont
formées, ce qui permet de conclure que su2a n’est pas actif sur m1.
Comme su1a est actif sur m1 et m2, alors que su2a n’est actif que sur m2, on peut imaginer :
– que su1a serait un suppresseur informationnel, allèle-spécifique et non gène-spécifique;
dans ce cas, les mutations m1 et m2 seraient deux mutations non-sens pour lesquelles le
suppresseur su1a apporterait un acide aminé compatible, alors que le su2a apporterait un
acide aminé compatible pour m2 mais pas compatible pour m1;
– que su1a serait un suppresseur physiologique pouvant agir sur m1 et m2; dans ce cas, peu
importe la nature des mutations m1 et m2, le suppresseur su2a étant éventuellement spécifique
de la mutation m2.
Compte tenu du caractère général des observations, il est inutile d’aller plus loin dans les
hypothèses explicatives.