Deux souches pures de drosophiles, A à yeux roses [r] et B à corps noir [n]
ont été croisées. Une F1 homogène de phénotype sauvage a été obtenue, à
yeux rouge brique et corps gris.
Le croisement F1 × F1 donne les résultats suivants en F2 : [+] 45 %;
[r] 30 %; [n] 25 %.
Interprétez ces résultats (nombre de gènes mis en cause, liaison physique
et génétique éventuelle, distances génétiques).
➤ Niveau Licence (L3)/Définition des objectifs.
– Analyse de la ségrégation et de la recombinaison génétique.
– Liaison physique et génétique.
Solution. Ce problème, qui paraît simple a priori, suppose en fait une très bonne maîtrise de
tous les concepts mis en oeuvre dans l’analyse génétique afin de poser, puis de résoudre les
différentes questions dans un enchaînement adéquat.
1. Le phénotype sauvage de la F1 permet de conclure que les phénotypes mutants sont récessifs,
vis à vis des phénotypes sauvages.
2. On supposera que les croisements réciproques ont été faits, ce qui exclut toute localisation
d’un des gènes sur le chromosome X.
3. Les deux souches croisées étant mutantes et de phénotype récessifs, on peut conclure
qu’elles sont forcément mutées dans au moins deux gènes différents, sinon l’absence de
complémentation fonctionnelle aurait conduit à un phénotype non sauvage de la F1 (on
exclut a priori l’hypothèse d’un gène muté avec deux mutations aux effets différents, l’une
sur la couleur de l’oeil, l’autre sur la couleur du corps pouvant conduire à une complémentation
fonctionnelle intragénique en F1).
Il y a donc au moins un gène muté impliqué dans la pigmentation du corps, et un autre gène
muté impliqué dans celle des yeux.
4. La question qui se pose alors est de savoir dans combien de gènes sont mutées chacune des
deux souches.
Pour cela, il suffit de tester la ségrégation allélique 2/2 à la méiose de la F1
pour chacun des phénotypes, sachant, compte tenu de la dominance, qu’une telle ségrégation
conduira à 3/4 de sauvages et 1/4 de mutés.
C’est le cas pour le phénotype de pigmentation du corps (75 % de « + » et 25 % de noirs) et
on peut conclure que la souche B est un mutant simple, muté dans un seul des gènes de
pigmentation du corps qui sera noté N (allèle muté n).
Ce n’est pas le cas pour le phénotype de coloration des yeux (70 % de « + » et 30 % de roses) et
on doit conclure que la souche A est mutée dans au moins deux gènes (mais pourquoi pas plus…).
Plusieurs questions se posent alors :
– la souche A est-elle mutée seulement dans deux gènes ou plus ?
– ayant dénombré les gènes mis en jeu, peut-on statuer sur leur indépendance génétique,
voire physique (voir les exercices précédents) ?
– en cas de liaison génétique, peut-on calculer leurs distances respectives ?
5. Nombre de gènes mutés chez A : il convient de partir d’une hypothèse minimaliste de
deux gènes mutés qui seront notés A et B (allèles mutés respectifs a et b).
Le génotype de la F1 est double hétérozygote et peut s’écrire (A … B) // (a … b) si on considère
que les pointillés figurent notre méconnaissance d’une éventuelle liaison entre les deux
gènes.
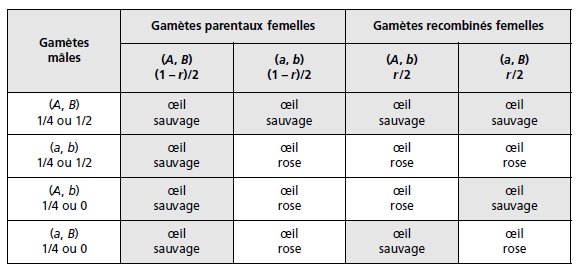
On sait qu’il n’y a pas de crossing-over chez la drosophile mâle, mais on va établir un tableau
de gamètes du croisement F1 × F1 (tabl. 3.28) dans la situation la plus large possible de deux
gènes physiquement indépendants, quitte à réduire ce tableau si les résultats attendus ne sont
pas conformes aux observations (voir exercice 3.4), en désignant par r le taux de recombinaison
entre les deux gènes (fréquence des gamètes recombinés).
On délaisse, dans cette
analyse, le gène N et ses phénotypes.
TABLEAU 3.28 TABLEAU DE CROISEMENT DES GAMÈTES POUR DEUX GÈNES.
r est le taux de recombinaison chez les femelles; chez les mâles les gamètes sont équifréquents en cas d’indépendance physique, les gamètes recombinés sont
absents en cas de liaison physique.
• Dans le cas de deux gènes physiquement indépendants, les seize cases de ce tableau sont
valides et équifréquentes (dans ce cas la valeur de r est égale à 1/2), et on attend 9/16
(56,25 %) de [sauvage], et 7/16 (43,75 %) de [oeil rose]; ce qui n’est pas le cas et exclut donc
l’hypothèse de deux gènes physiquement indépendants.
• Dans le cas de deux gènes physiquement liés, mais génétiquement indépendants, les
gamètes mâles sont exclusivement parentaux et les deux dernières lignes du tableau ne
doivent pas être prises en compte.
Comme r est ici égal à 1/2, on attend 5/8 (62,5 %) de [oeil sauvage] et 3/8 (37,5 %) de [oeil
rose], ce qui n’est pas le cas et exclut donc cette hypothèse.
• Dans le cas d’une liaison physique et génétique (valeur de r inférieure à 1/2), on observera
alors les phénotypes avec les fréquences suivantes, obtenues par sommation des fréquences
des génotypes sous-jacents :
f [oeil sauvage] = 1/2 + (1 – r)/4; f [oeil rose] = r/2 + (1 – r)/4.
Comme f [oeil rose] = 0,3, on en déduit facilement la valeur r = 0,2.
Les observations sont cohérentes avec l’hypothèse minimaliste de deux gènes A et B distants
de 20 ur.
Remarque. Cette analyse a été développée sous une hypothèse fonctionnelle implicite
que les gènes A et B étaient « complémentaires », à savoir qu’il suffit d’être a//a ou b//b
pour avoir le même phénotype mutant (exercice 3.5).
On peut vérifier que l’hypothèse
de deux gènes redondants (exercice 3.6; seul le double homozygote récessif est de
phénotype muté) est incohérente avec les observations.
6. Il convient maintenant de situer le gène N par rapport aux deux gènes A et B, sachant qu’il
sont liés, ce qui va limiter nos catégories de gamètes chez les mâles, mais non chez les
femelles qui peuvent, avec un génotype triple hétérozygote, former 8 types de gamètes.
Le génotype de la F1, pour les trois gènes N, A et B, est triple hétérozygote et peut s’écrire
(n … A-B)//(N … a-b) si on considère que les pointillés figurent notre méconnaissance d’une
éventuelle liaison entre le gène N et le bloc A-B; sans savoir, en cas de liaison, lequel de A, B
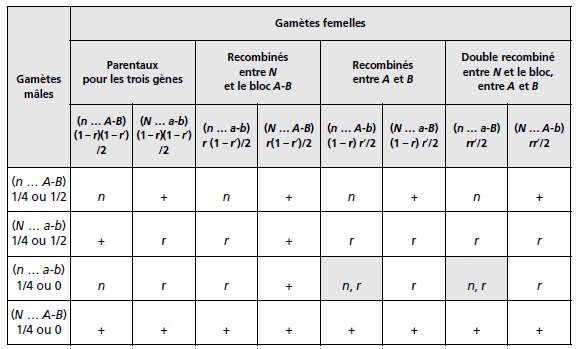
ou N est central. On peut alors construire la F2 issue d’un croisement F1 × F1 par un tableau
de croisement des gamètes (tabl. 3.29).
On note, pour simplifier, les seuls phénotypes mutés
et les sauvages (voir énoncé).
TABLEAU 3.29 GÉNOTYPES ET PHÉNOTYPES RÉSULTANT DE L’UNION DES GAMÈTES
À L’ISSUE D’UN CROISEMENT F1 × F1.
r est le taux de recombinaison entre N et le bloc A-B et r′ celui entre A et B
(r′ = 0.2), chez les femelles. Chez les mâles les gamètes sont équifréquents en cas
d’indépendance physique, les gamètes recombinés sont absents en cas de liaison
physique.
• En cas d’indépendance physique de N, les 32 cases du tableau sont valides et on devrait
observer des phénotypes double mutés [noir, rose], ce qui n’est pas le cas et exclut cette
hypothèse.
Il y a donc liaison physique des trois gènes.
• Puisqu’il y a liaison physique, la question se pose de savoir si N est central (entre A et B)
ou extérieur au segment A-B.
Le tableau 3.29 a été construit sous cette deuxième hypothèse
(voir fréquence des gamètes double recombiné); il permet, en ne considérant que les deux
premières lignes, seules valides pour les gamètes mâles, de calculer les fréquences des trois
phénotypes, en sommant celles des génotypes sous-jacents, soit :
L’avantage de ces formules algébriques est de pouvoir montrer immédiatement, et sans ambiguïté,
que les fréquences sont ici indépendantes de la valeur de r et ne dépendent que de la
valeur de r′.
Quelle que soit la valeur de r′, la fréquence f [n] est égale à 1/4 et correspond à la ségrégation
du couple d’allèles N/n, la fréquence des sauvages pour le phénotype de couleur du corps
obtenu égale à f [+] + f [r] = 3/4.
Les fréquences f [+] et f [r] ne dépendant que du taux de recombinaison entre A et B sont obligatoirement
égales à 0,45 et 0,30 et ne peuvent pas permettre de situer N par rapport au bloc
A-B (liaison ou indépendance génétique), dans l’hypothèse où il serait extérieur au segment
A-B.
Il reste à traiter l’hypothèse où N serait situé entre A et B.
Dans le cas où N est central, le tableau de croisement des gamètes comporte les mêmes lignes
et les mêmes colonnes, mais il convient de modifier les fréquences des gamètes :
• en prenant r fréquence de recombinaison entre A et N et r′ fréquence de recombinaison
entre N et B;
• en considérant que les gamètes doubles recombinés sont dans ce cas A-N-B et a-n-b
(fréquence r * r′/2).
Les sommes algébriques sont alors égales à
f[+]= (2-r′) + (2rr′)/4=(2-R)/4
f[r]= (2+r+r′) - (2rr′)/4=(2+R)/4
f[n]=1/4
où R = r + r′ – 2rr′, le taux de recombinaison entre A et B (voir page 53).
Au bout du compte les informations apportées par le croisement étudié permettent de
montrer la liaison physique de N avec A et B mais laissent indéterminée sa position centrale
ou externe, et dans ce dernier cas, son éventuelle indépendance génétique.