1. • Analyse génétique. R1 à R5 sont des révertants, capables d’utiliser le lactate, et donc le
glycérol.
Comme le mutant de départ est délété pour le gène de l’iso1-cytochrome c, ces
révertants sont obligatoirement porteurs d’un suppresseur physiologique, noté sua, dont
l’effet est de rendre inutile le produit codé par le gène muté, ici cyc-1.
Ici le suppresseur
physiologique a pour effet d’accroître la quantité d’iso2-cytochrome c de façon telle, audessus
du 5 % sauvage, que la croissance sur lactate devient possible.
• Interprétation fonctionnelle et biochimique.
L’accroissement de la quantité d’iso2-cytochrome
c peut résulter de plusieurs événements moléculaires distincts :
– ce peut être une mutation affectant l’allèle sauvage cyp3+, soit son promoteur qui serait
plus actif, par exemple en ayant plus d’affinité pour l’activateur de CYP3 (on sait qu’il y a
une régulation positive), soit la séquence codante qui donnerait une molécule plus active
(hypothèse peu vraisemblable mais pas impossible);
– ce peut être une mutation affectant le gène de l’activateur de CYP3, le rendant plus actif.
2. Les diploïdes obtenus ont respectivement les génotypes (on note sua, la séquence mutée
ayant un effet suppresseur actif, et sui, la séquence sauvage inactive) :
Que le premier soit sauvage [gly+, lct+] est logique, puisqu’on sait que cyc1+ est dominant et
que sua a un effet suppresseur sur celui de cyc1-1.
Mais le fait que le second soit également
sauvage [gly+, lct+] prouve que l’effet de sua est dominant sur celui de sui.
L’analyse de tétrades issues des méioses des diploïdes R1 × S′ teste la ségrégation aux
locus cyc1 et su.
On peut attendre quatre types de spores de génotype et de phénotype :
– parental (cyc1-1, sua), donc de phénotype [gly+, lct+];
– parental (cyc1+, sui), donc de phénotype [gly+, lct+];
– recombiné (cyc1-1, sui), donc de phénotype [gly+, lct–];
– recombiné (cyc1+, sua), a priori de phénotype [gly+, lct+], mais cela doit être démontré.
Les 20 tétrades avec 4 spores [gly+, lct+] sont des DP, et les deux autres types de tétrades sont
des DR et des T.
Comme les DR contiennent au moins deux spores recombinées [gly+, lct–],
on peut en déduire que les deux autres spores recombinées de ces 19 DR sont de phénotype
[gly+, lct+], comme attendu.
Les 21 autres sont des T.
Les gènes cyc1 et su sont génétiquement indépendants, puisque les fréquences des DP et des
DR sont égales, et physiquement indépendants, puisque la fréquence des T est inférieure à 2/3.
L’analyse de tétrades issues des méioses des diploïdes R1 × A′ teste la ségrégation 2/2 pour
le suppresseur, on doit en effet attendre deux types de spores équifréquentes, de génotypes
sua ou sui et de phénotype [gly+, lct+] ou [gly+, lct–] et un seul type de tétrade, si l’effet
suppresseur résulte d’une seule mutation, ce qui est confirmé par le résultat.
3. Les diploïdes obtenus ont respectivement les génotypes (on note par su1a et su3a, les
suppresseurs actifs de R1 et R3) :
R1 × C : (cyc1-1, cyp3+, su1a)//(cyc1-1, cyp3-10, su1i);
R3 × C : (cyc1-1, cyp3+, su3a)//(cyc1-1, cyp3-10, su3i).
Que ces diploïdes soient sauvages [gly+, lct+] est logique, la perte de fonction de cyp3-10 est
récessive et compensée par l’effet de cyp3+, et on sait que su1a ou su3a ont un effet suppresseur
dominant vis-à-vis de l’effet ou l’absence d’effet de su1i ou su3i.
L’analyse de tétrades issues des méioses des diploïdes R1 × C teste la ségrégation aux
locus cyp3 et su1, afin de tester la liaison ou l’indépendance génétique.
On peut attendre
quatre types de spores de génotype et de phénotype :
– parental (cyp3+, su1a), donc de phénotype [gly+, lct+];
– parental (cyp3-10, su1i), donc de phénotype [gly–, lct–];
– recombiné (cyp3+, su1i), donc de phénotype [gly+, lct–];
– recombiné (cyp3-10, su1a), a priori de phénotype [gly–, lct–], puisque les deux gènes sont
mutés et que l’effet de su1a d’accroître l’expression de la séquence CYP3 ne doit pas être
« très suppresseur », si cette séquence est mutée (cyp3-10), mais cela doit être démontré.
Les 6 tétrades avec 2 spores [gly+, lct+] et 2 spores [gly–, lct–] sont obligatoirement les DP, et
les deux autres types de tétrades sont des DR et des T.
Comme les DR contiennent au moins
deux spores recombinées [gly+, lct–], on peut en déduire que les deux autres spores recombinées
de ces 7 DR sont de phénotype [gly–, lct–], comme attendu. Les 17 autres sont des T.
Les gènes CYP3 et su1 sont génétiquement indépendants, puisque les fréquences des DP et
des DR sont égales, mais on ne peut statuer sur l’indépendance physique, puisque la
fréquence des T est égale à 2/3.
L’analyse de tétrades issues des méioses des diploïdes R3 × C teste la ségrégation aux locus
CYP3 et su3, qui n’est pas obligatoirement le même que su1, afin de tester la liaison ou l’indépendance
génétique.
On peut attendre quatre types de spores de génotype et de phénotype :
– parental (cyp3+, su3a), donc de phénotype [gly+, lct+];
– parental (cyp3-10, su3i), donc de phénotype [gly–, lct–];
– recombiné (cyp3+, su3i), donc de phénotype [gly+, lct–];
– recombiné (cyp3-10, su3a), a priori de phénotype [gly–, lct–], puisque les deux gènes sont
mutés et que l’effet de su3a d’accroître l’expression de la séquence mutée cyp3-10 ne doit
pas être « très suppresseur », mais cela doit être démontré.
Les 85 tétrades avec 2 spores [gly+, lct+] et 2 spores [gly–, lct–] sont obligatoirement les DP,
et l’autre tétrade ne peut être qu’un T, car il y aurait deux spores [gly+, lct–] si c’était un DR;
les deux spores [gly–, lct–] correspondent bien alors au parental et au recombiné attendu.
D’ailleurs, en cas de liaison génétique, comme on va le voir, la fréquence des DR (qui supposent
au moins 2 crossing-over) est toujours inférieure à celle des T (qui n’en supposent qu’un
seul).
Les gènes CYP3 et su3 sont génétiquement très liés, puisque la fréquence des DP est très
supérieure à celle des DR et la distance entre les locus est égale à d = [3f(DR) + f(T)/2] × 100
= 0,58 ur.
Si on confronte les résultats relatifs à R1 et R3 aux hypothèses biochimiques détaillées à la
question 1, on peut admettre que R1 n’est certainement pas muté dans l’opérateur de CYP3+
et qu’il l’est sans doute dans son activateur, et que R3 pourrait être muté dans l’opérateur de
CYP3+, compte tenu de la distance très faible entre les sites su3a et cyp3-10; cependant la
distance n’est pas un critère absolu, car on pourrait imaginer que su3a est muté dans un gène
contigu de CYP3.
4. a Il est logique de prendre la tétrade T, puisqu’on est sûr d’avoir, parmi les deux spores
[gly–, lct–], la spore recombinée de génotype (cyc1-1, cyp3-10_ su3a) tandis que l’autre est le
parental de génotype (cyc1-1, cyp3-10_ su3i).
NB : le signe « _ » entre les locus cyp3 et su3 indique la liaison génétique.
Les diploïdes obtenus par croisements parallèles avec A ou A′ sont de génotypes :
(cyc1-1, cyp3-10_su3a)//(cyc1-1, cyp3+_sui3i)
et (cyc1-1, cyp3-10_ sui3i)//(cyc1-1, cyp3+_ sui3i).
Que le deuxième génotype soit de phénotype [gly+, lct–] est attendu puisqu’il n’a qu’une
copie sauvage de l’allèle cyp3+ et ne peut, au mieux, que pousser sur glycérol; mais que le
premier génotype soit aussi de phénotype [gly+, lct–], alors que su3a est présent, prouve que
la séquence su3a n’a pas d’effet sur cyp3+, chez ce diploïde, alors qu’elle avait un effet sur
cyp3+ chez le révertant R3.
Cela prouve que su3a est cis-dominant et qu’il affecte, sans doute, le promoteur de CYP3,
hypothèse d’autant plus vraisemblable sachant la distance génétique entre les sites su3a
et cyp3-10.
4. b Le révertant R3 correspond bien à une des deux hypothèses biochimiques de la
question 1, et la cis-dominance est la propriété attendue du mutant de cible dont on a fait
l’hypothèse à la question 3.
4. c Si su3a n’avait pas été cis-dominant mais trans-dominant, affectant le gène d’un activateur,
par exemple, alors le premier génotype diploïde aurait été de phénotype [gly+, lct+].
5. D et E restent dépourvus d’iso1 parce qu’ils sont restés cyc1-1 !
Si E est dépourvu d’iso2, on peut supposer qu’il est muté dans la séquence codante de CYP3,
ou bien dans le gène de l’activateur de CYP3, c’est-à-dire dans le gène su1, si on suppose que
su1a affectait le gène de l’activateur.
En revanche, D présentant encore une activité même réduite d’iso2, ne peut être muté dans la
séquence codante de CYP3, mais peut l’être soit dans son promoteur, soit, là encore, dans le
gène de l’activateur, l’activité iso2 réduite correspondant au taux de transcription de base du
gène CYP3+ en absence d’activateur.
Mais on peut aussi imaginer que D et/ou E puissent être mutées dans un autre gène que cyp3
ou son activateur !
Aussi les génotypes des diploïdes obtenus par croisement parallèles de D et E avec R1′
peuvent être écrits formellement ainsi :
(cyc1-1, cyp3+, d, su1a)//(cyc1-1, cyp3+, d+, su1a)
et (cyc1-1, cyp3+, e, su1a)//(cyc1-1, cyp3+, e+, su1a).
où d et e sont les « notations formelles » des mutations affectant respectivement D et E, les
mutations d ou e pouvant affecter le gène CYP3 (dans ce cas une des copies est cyp3–) ou le
gène de l’activateur (dans ce cas une des copies est sui1) ou un autre gène.
Il est logique que les phénotypes soient dans les deux cas [gly+, lct+] puisque au moins un des
deux su1a est fonctionnel, mais cela montre aussi que les mutations d et e sont récessives.
L’analyse de ségrégation montre que D ou E ne diffèrent de R1 que pour un seul gène (ségrégation
2/2 des couples d//d+ ou e//e+); ce gène étant donc CYP3 ou sua1 (activateur de CYP3)
ou un troisième.
Les génotypes des diploïdes obtenus par croisements parallèles de D et E avec C peuvent être
écrits ainsi :
D × C : (cyc1-1, cyp3+, d, su1a)//(cyc1-1, cyp3-10, d+, su1i)
et E × C : (cyc1-1, cyp3+, e, su1a)//(cyc1-1, cyp3-10, e+, su1i).
• Le premier diploïde est de phénotype [gly+, lct–], ce qui est possible si la mutation affecte
le gène de l’activateur et que la séquence su1a disparaît au profit d’une perte de fonction
notée su–, le gène CYP3 n’étant plus régulé que par la copie sauvage su1i, ce qui redonne le
phénotype [gly+, lct–].
Chez l’haploïde D, la mutation su– abaisse le taux de transcription de CYP3+ à un « niveau de
base » donnant une quantité d’iso2 insuffisante pour pousser sur glycérol, mais dosable
(2 % de la quantité SSR).
• Le deuxième diploïde est de phénotype [gly–, lct–], ce qui est possible si la mutation e
touche l’allèle cyp3+ qui devient une mutation de perte de fonction cyp3–, il n’y a alors plus
aucun cytochrome c, ni chez E (comme cela a été observé, dosage = 0) ni chez le diploïde.
Toutefois, sur un plan formel, rien n’exclut que les mutations d ou e touchent un troisième
gène, même si on montrait que d coségrège avec su1 et que e coségrège avec CYP3.
Problème 10.8
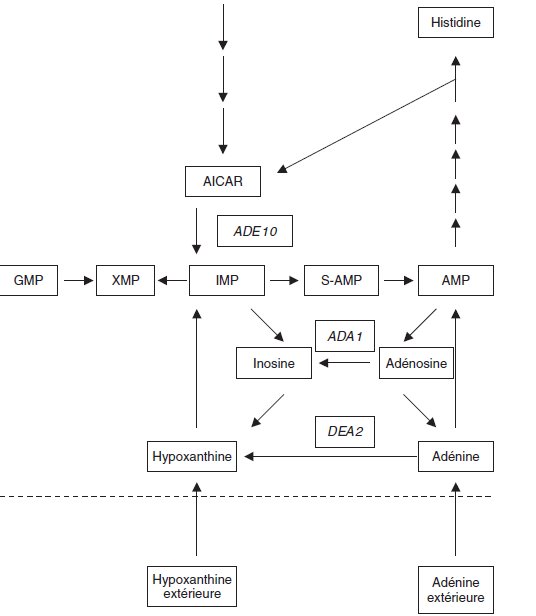
Le schéma ci-contre présente une partie du métabolisme de l’adénine et de
l’hypoxanthine (dont la formule diffère de celle de l’adénine par l’absence de
fonction amine sur le carbone 6 du cycle, remplacée par une fonction
cétone) chez la levure Saccharomyces cerevisiae.
L’inosine est le nucléoside de l’hypoxanthine comme l’adénosine est celui
de l’adénine; à partir de l’IMP (inosine monophosphate) la cellule réalise
la synthèse de xanthine monophosphate puis de GMP (guanosine monophosphate).
La cellule a donc besoin, pour se développer, de pouvoir assurer à la fois la
synthèse d’adénine et d’hypoxanthine.
Question 1.
Le gène ADE 10 spécifie l’enzyme qui assure la transformation de AICAR
en IMP. Ce gène a été identifié chez un mutant haploïde auxotrophe ade10 de phénotype [ade–];
Justifiez ce phénotype et le fait que le mutant ade10
peut aussi bien pousser sur un milieu minimum (Mo) supplémenté en adénine
qu’en hypoxanthine.