C’est un mécanisme de transfert de gènes d’une bactérie donatrice à une bactérie
réceptrice, via une capside virale.
Certains bactériophages (nom donné au virus bactérien) comme P22 chez Salmonella
ou P1 chez coli induisent, durant leur cycle lytique, une fragmentation du
génome bactérien qui conduit certains de ces fragments à une encapsidation à la
place d’un génome viral (environ 100 000 pb).
Ces phages dits transducteurs sont alors capables, en infectant des bactéries
réceptrices, de leur transférer ce fragment de génome bactérien qui peut alors
remplacer, par recombinaison homologue, une partie ou la totalité de la séquence homologue endogène.
Ainsi, une souche de bactéries auxotrophes pour la valine
traitée avec un lysat transducteur préparé sur une souche sauvage verra l’apparition
de recombinants sauvages [val+], identifiables par étalement des réceptrices survivantes
sur milieu minimum et récupération de colonies [val+].
Bien sûr, la multiplicité d’infection (nombre moyen de phages par bactérie) doit
être très inférieure à 1 car une infection multiple conduirait toutes les réceptrices, notamment
celles infectées par un phage transducteur, à être aussi infectées par un « vrai »
phage dont l’effet est lytique.
Les bactéries ne doivent jamais être infectées par plus d’un
phage afin de laisser survivre les recombinants issus de l’infection par un phage transducteur
apportant la séquence homologue de la séquence endogène à recombiner.
La transduction est un moyen puissant de cartographie des gènes et même de
cartographie fine (sites de mutations très proches, voire intragéniques).
Si, par transduction,
une souche porteuse de plusieurs mutations peut être recombinée pour
toutes celles-ci, cela prouve qu’il y a eu cotransduction (la multiplicité d’infection
est inférieure à 1) et que tous les sites de mutations sont localisés sur un fragment de
génome dont la taille maximale est égale au génome viral, soit environ 100 000 pb.
La cotransduction permet, en fonction des génomes donneurs et receveurs, et des
fréquences des différents recombinants, de réaliser un « test trois points » et de
définir entre trois sites lequel est central.
La transduction définie ici est la transduction généralisée; la « transduction
spécialisée » qui n’affecte que quelques gènes bactériens n’est pas abordée dans cet
ouvrage.
NB : La transduction est aussi un outil efficace pour construire des souches par
transfert de mutations de l’une à l’autre.
C - La transformation
:
Il fallut attendre 1943 pour que Avery et McLeod, puis d’autres bactériologistes,
montrent que la transformation bactérienne observée par Griffiths en 1928 chez
Streptococcus pneumoniae résultait simplement de la capacité d’une souche bactérienne
à laisser entrer de l’ADN exogène (on dit exogénote) nu susceptible de venir
transformer le génome endogène (on dit endogénote) par recombinaison, ou de le
compléter par addition, dans le cas d’un plasmide.
La transformation suppose, dans les conditions naturelles, que les bactéries réceptrices
soient « compétentes », un état physiologique permettant une entrée passive
ou active de l’ADN exogène (le mécanisme est différent selon que les bactéries sont
gram+ ou gram–).
La taille de l’ADN exogène impliqué dans la transformation ne peut guère dépasser
10 000 pb et ne peut donc impliquer autant de gènes que les fragments opérant dans
la transduction.
Par ailleurs, la transformation est beaucoup moins efficace que la
transduction, du fait que les réceptrices doivent être compétentes tandis qu’elles sont
toujours, sauf exception, aptes à être infectées par un phage transducteur.
La découverte de la conjugaison bactérienne et de la transduction ont fortement
limité l’intérêt de la transformation comme outil d’analyse génétique.
En revanche, la transformation bactérienne est un outil indispensable du génie
génétique et de la biologie moléculaire.
La transformation d’une réceptrice par un
plasmide est devenue une opération courante pour constituer des banques génomiques,
cloner un gène (tri de la banque, clonage direct…), le séquencer, étudier son
expression, réaliser une mutagenèse ciblée de celui-ci, etc.
De nombreuses opérations de la biologie moléculaire sont faciles à réaliser et de
rendement efficace chez la bactérie.
Les plasmides sont un excellent vecteur moléculaire
pour toutes les manipulations de l’ADN, ils sont faciles à purifier, à manipuler
ou à traiter in vitro, à réinsérer par transformation dans une bactérie qui
assurera le clonage amplificateur de la séquence portée par le plasmide.
Remarque 1. Lors de la transformation d’une bactérie par un plasmide, en
général porteur d’un gène de sélection positive comme la résistance à un antibiotique,
l’ADN du plasmide constitue un réplicon qui reste indépendant du
génome, un peu comme l’épisome.
De ce fait, on peut aussi, en clonant de
l’ADN bactérien dans un plasmide, obtenir des mérodiploïdes susceptibles de
permettre les tests de l’analyse génétique, dominance et complémentation
fonctionnelle, mais il ne faut pas oublier que le fragment cloné est toujours
d’une dimension restreinte, la transformation ne pouvant impliquer un ADN
de taille supérieure à 10 000 pb.
Remarque 2. La linéarisation d’un plasmide lui fait perdre le statut de
réplicon, et la transformation d’une réceptrice par un plasmide linéarisé ne
peut conduire qu’à sa perte ou à l’intégration de tout ou partie du plasmide par
recombinaison homologue à partir d’une des séquences de son extrémité (on
cible donc l’insertion d’un plasmide dans le génome récepteur en le linéarisant
à un endroit ou un autre).
Exercices
:
Exercice 9.1
On dispose d’une souche H d’Escherichia coli, Hfr, prototrophe, sensible à
la streptomycine, phénotype noté [strS], et au phage T6, phénotype noté
[tsxS].
On dispose d’une souche A d’Escherichia coli, F–, auxotrophe pour la
thréonine, phénotype noté [thr–], incapable de métaboliser le galactose,
phénotype noté [gal–], résistante à la streptomycine, phénotype noté [strr],
et au phage T6, phénotype noté [tsxr].
Le phage T6 est comparable au phage T4, sa virulence entraîne, dès
l’infection de la bactérie, un détournement vers le cycle lytique des
composés bactériens accompagnés de dérèglements métaboliques et de
lésions immédiates de l’ADN bactérien.
On effectue le croisement (coculture) H × A.
• Dans une première expérience (tabl. 9.1), on prélève, à des temps variables,
des échantillons d’un même volume de la culture que l’on dilue puis
qu’on étale sur un milieu gélosé contenant de la streptomycine et, soit du
glucose, soit du galactose avec de la thréonine.
On compte alors le nombre
de colonies obtenues sur chacun des deux milieux aux différents temps
d’étalement.
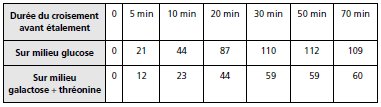
TABLEAU 9.1.
• Dans une deuxième expérience, on prélève, à des temps variables, des
échantillons d’un même volume de la culture, auxquels on ajoute une
quantité saturante de phage T6 avant dilution et étalement sur un milieu
gélosé contenant de la streptomycine et, soit du glucose, soit du galactose
avec de la thréonine. On compte alors le nombre de colonies obtenues sur
chacun des deux milieux aux différents temps d’étalement.
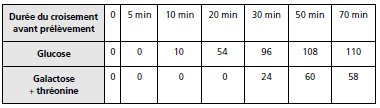
TABLEAU 9.2.
1. Interprétez la deuxième expérience :
– en précisant le rôle du phage T6. Comment feriez-vous l’étude dans le
cas où votre stock de phage T6 serait épuisé ?
– en concluant sur le plan cartographique.
2. En quoi et pourquoi les résultats de la première expérience diffèrent-ils
de ceux de la seconde, sans leur être contradictoires ? Que permettent-ils
de conclure ? Quelle précision ne peuvent-ils apporter ?
3. Pourquoi le nombre de colonies [gal+] ou [thr+] finit par atteindre un
maximum ? Pourquoi ce maximum n’est pas le même pour les deux
phénotypes recombinants ?
– Comprendre la conjugaison et ses conséquences génétiques.
– Cartographie des gènes par conjugaison.
Solution
1. En ajoutant le phage T6 on tue les bactéries sensibles de la souche Hfr et on interrompt la
conjugaison puisqu’il s’agit d’un phénomène actif où l’ADN est dupliqué puis transféré vers
la réceptrice.
Les réceptrices sont résistantes et échappent à l’action de T6 et pourront se développer dans
le milieu d’étalement, du fait de leur résistance à la streptomycine, mais à la condition d’avoir
reçu et recombiné la séquence sauvage homologue de la mutation thr–, pour le premier
milieu, ou la séquence sauvage homologue de la séquence mutée gal– pour le second milieu.
Ce protocole de conjugaison interrompue permet de conclure que la séquence thr+ n’est pas
transférée avant au moins 5 min de conjugaison (entre 5 et 10) et que la séquence gal+ ne
l’est pas au moins avant 20 min (entre 20 et 30); les deux mutations sont donc distantes
d’environ 15 min de temps de conjugaison.
En absence de phage T6, on procéderait à une forte agitation (vortex) qui rompt les ponts
cytoplasmiques.
Remarque.
La précision d’un protocole de conjugaison interrompue peut descendre
facilement à un intervalle d’une minute; par ailleurs, en faisant un graphe et en extrapolant
vers l’axe des abscisses la droite d’accroissement du nombre de colonies, on
peut estimer plus finement le temps de conjugaison minimal avant l’entrée de chacun
des gènes.
2. Dans la première expérience, des colonies [thr+] et même [gal+] apparaissent dès la
cinquième minute alors qu’on vient de conclure que les séquences thr+ et gal+ n’entraient pas
avant 5 et 20 min respectivement, mais ces observations n’ont qu’une contradiction apparente
avec celles de la deuxième expérience, car ici l’absence de traitement au phage T6
(et d’agitation) laisse les conjugaisons se poursuivre sur la boîte après l’étalement.
Ce protocole de conjugaison non interrompue est beaucoup moins précis car il donne l’ordre
d’entrée mais pas la distance en temps de conjugaison.
Seule l’interruption permet d’affirmer
qu’avant un temps donné la séquence testée n’est jamais entrée.
Dans la mesure où il y a toujours plus de thr+ que de gal+, on doit conclure que thr+ passe en
premier.
En effet si tel est le cas, il y aura un certain nombre de conjugaisons spontanément
interrompues entre le passage de thr+ et celui de gal+, de sorte que le nombre de réceptrices
ayant reçu thr+ sera toujours plus élevé que le nombre de réceptrices ayant reçu gal+.
3. La différence de fréquence entre les deux types de recombinants vient d’être explicitée; il
reste à comprendre pourquoi la fréquence de chaque type recombinant tend vers un maximum.
En fait, la population de réceptrices est une population finie; pour chaque réceptrice conjugant,
il existe une probabilité d’interruption spontanée après le transfert d’un gène donné;
probabilité dont la valeur est d’autant plus grande que ce gène est près de l’origine de transfert.
De ce fait, il y aura un maximum de recombinants, pour ce gène, égal au produit du
nombre de réceptrices par la probabilité d’interruption après le transfert du gène.
Ce
maximum sera atteint dès que le temps de culture est tel que presque toutes les réceptrices
ont commencé à conjuguer.
Ce maximum est fonction de la probabilité d’interruption après le passage du gène et donc de
la proximité du gène par rapport à l’origine de transfert.
Le niveau des « plateaux » ou des
« plafonds » donne bien l’ordre d’entrée des gènes.