Ce chapitre se borne à des rappels et à un résumé des notions principales de génétique
bactérienne, et concerne exclusivement les eubactéries qui, comme le colibacille
(Escherichia coli), le pneumocoque ou la salmonelle, renferment un
chromosome constitué d’une molécule d’ADN double-brin, nu et circulaire.
Introduction :
La génétique est née dans le monde eucaryote, le fondement de son analyse repose
sur l’observation des diploïdes issus de croisements
conçus par l’expérimentateur (test de dominance ou test de complémentation fonctionnelle)
puis l’étude des produits de la méiose chez ces diploïdes (test de la ségrégation
2/2 ou de la liaison génétique).
Par l’ensemble de ce dispositif expérimental, le généticien peut, à partir de
plusieurs mutants indépendants, dont le phénotype diffère d’un phénotype de référence,
déterminer si ce sont des mutants simples ou multiples, lesquels sont mutés
dans le même ou un même gène, dénombrer ainsi le nombre minimal de gènes
impliqués dans le phénotype ou le phénomène biologique étudié, cartographier les
gènes liés, voire les sites de mutations au sein d’un même gène.
Or aucun de ces principes expérimentaux, observation des diploïdes ou analyse
des produits de leurs méioses, ne peut s’appliquer à l’analyse génétique chez les procaryotes pour une raison simple et évidente : les procaryotes, et, parmi eux, les
eubactéries comme Escherichia coli, ne sont jamais diploïdes.
Aussi la génétique bactérienne se fonde sur des propriétés spécifiques des bactéries
pour entreprendre sa démarche analytique.
Celle-ci est utile parce que des mécanismes
fondamentaux à tout le monde vivant y sont souvent plus simples à étudier
que chez la souris ou la drosophile, mais aussi parce que la variété extrême du
monde bactérien est une mine de découvertes, pour la biologie fondamentale et pour
les biotechnologies du futur.
Toute étude génétique d’un phénomène suppose d’en voir des variants, ce qui est
assez facile chez les bactéries qui, comme tous les organismes unicellulaires,
peuvent être facilement cultivées dans un milieu simple (milieu minimum) liquide
ou solide, au sein duquel on peut cribler des mutants du métabolisme, de résistance
à des toxiques, ou mutés dans des fonctions cellulaires plus essentielles.
Mécanismes bactériens de substitution ou de complément de
l'information génétique endogène :
La génétique bactérienne se fonde sur trois phénomènes ou mécanismes naturels
permettant, chez les bactéries, l’entrée d’ADN exogène venant compléter ou
remplacer localement l’information endogène. Ces trois phénomènes sont la conjugaison,
la transduction et la transformation.
A - La conjugaison
:
La conjugaison bactérienne a été découverte en 1946 par Josuah Lederberg et
Edouard Tatum chez E. coli.
Ils démontrèrent sans ambiguité que deux souches
bactériennes porteuses de nombreuses mutations d’auxotrophie différentes
pouvaient, lors d’une coculture (croisement bactérien), donner des recombinants
prototrophes capables de pousser sur une boîte de milieu minimum, contrairement
aux deux souches parentales.
Ils établirent la nécessité d’un contact entre bactéries
(excluant ainsi l’hypothèse de la transformation, voir plus loin) et visualisèrent par
microscopie l’établissement d’un pont cytoplasmique à travers lequel on pouvait
supposer un échange d’ADN, une bactérie réceptrice recevant les séquences
sauvages d’une bactérie donatrice puis remplaçant par celles-ci les séquences
mutées endogènes, acquérant ainsi un génotype sauvage.
Le nombre de mutations en
jeu dans les souches parentales et la fréquence des recombinants sauvages excluait
tout autre phénomène comme des mutations reverses ou suppressives.
On a introduit le terme de « parasexualité » pour rendre compte du phénomène de
conjugaison bactérienne et de ses conséquences génétiques, puisqu’elle permet le
brassage des gènes, comme la sexualité chez les eucaryotes.
Hayes, en 1953, établit que le transfert de gène est toujours unilatéral; des deux
souches en co-culture l’une est donneuse et l’autre réceptrice. Les seuls recombinants
sauvages sont des bactéries de la souche réceptrice ayant reçu, des bactéries de
la souche donatrice, les séquences sauvages venant remplacer les séquences mutées
endogènes; aucun recombinant sauvage ne pouvant être issu de bactéries de la
souche donatrice.
Pour rendre compte de ce statut donatrice/réceptrice et de la polarité du transfert
de gènes, Hayes a postulé puis démontré l’existence d’un facteur de fertilité F dont
la donatrice est pourvue et la réceptrice dépourvue; la première est ainsi désignée F+
et la seconde F–.
Cependant le facteur F, bien qu’étant une information génétique, se comportait
différemment des gènes.
Si les gènes étaient transmis avec une fréquence d’environ
10–7, le facteur F était transmis de façon infectieuse.
Dans un croisement, la plupart
des bactéries de la souche réceptrice devenaient F+, et restaient mutées, auxotrophes,
tandis que les rares recombinants, qui devenaient sauvages, restaient F–.
Ce paradoxe trouva sa solution avec la découverte par Cavalli-Sforza de bactéries Hfr (High frequency of recombination).
Les Hfr, contrairement aux bactéries F+, ne
transmettaient plus leur facteur F de fertilité mais transféraient leurs gènes 1 000 fois
plus efficacement, d’où un taux moyen de recombinants sauvages de 10–4 contre 10–7.
En fait, les bactéries d’une souche F+ possèdent en plus de leur chromosome
(attaché à la membrane plasmique) un « épisome », molécule d’ADN double-brin,
nue et circulaire mais libre.
L’épisome F (environ 100 Kb) est 10 à 20 fois plus grand
qu’un plasmide et se réplique de façon autonome (c’est un réplicon).
Il contient une
centaine de gènes dont ceux qui permettent l’établissement d’un pont cytoplasmique
dans l’acte « parasexuel » avec une réceptrice, puis le transfert infectieux d’une
copie de lui-même à celle-ci qui devient alors F+ (fig. 9.1).
Ainsi, dans la conjugaison entre bactéries F+ et F–, seul l’épisome est transféré, ce
qui explique son caractère infectieux, alors qu’aucun gène chromosomique n’est
transféré.
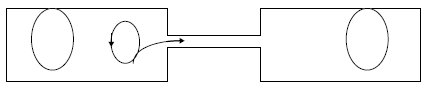
Figure 9.1 Le pont cytoplasmique entre la bactérie donatrice (à gauche)
et la bactérie réceptrice (à droite).
Il permet le passage d’une copie de l’épisome F; le transfert a toujours lieu à partir
d’une séquence spécifique nommée origine de transfert (en gras).
Par recombinaison homologue en un certain nombre de sites répartis sur le chromosome
bactérien, l’épisome F peut s’intégrer à celui-ci.
La bactérie devient alors Hfr car la conjugaison bactérienne, pilotée par l’épisome F, va conduire celui-ci, parce qu’il est intégré au chromosome bactérien, à entraîner avec lui tous les gènes
chromosomiques qui sont physiquement attachés derrière son origine de transfert
(fig. 9.2).
Si la conjugaison dure assez longtemps (90 à 100 minutes) sans être interrompue,
une copie de la totalité du chromosome bactérien peut être transférée.
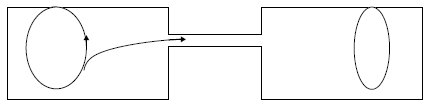
Figure 9.2 Le pont cytoplasmique entre la bactérie donatrice (à gauche)
et la bactérie réceptrice (à droite). Il permet le passage d’une copie de l’épisome F et, avec elle, une copie du chromosome,
partielle ou totale selon la durée de la conjugaison. Le transfert a toujours
lieu à partir de la séquence spécifique d’origine de transfert de l’épisome (en gras).
En fonction du lieu et du sens d’insertion de l’épisome, c’est-à-dire en fonction de
la souche Hfr, ce ne sont pas les mêmes gènes qui sont transférés à la suite immédiate
de l’origine de transfert (fig. 9.3).
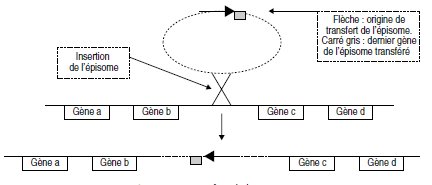
Figure 9.3 Transfert de l’épisome.
Ainsi inséré, le transfert de l’épisome, à partir de sa séquence d’origine de transfert,
entraînera les gènes c et d en premier et les gènes a et b en dernier, s’il n’y a pas
d’interruption du transfert (inséré dans l’autre orientation, l’épisome entraînerait
d’abord b et a, puis d et c en dernier). On remarque alors que la totalité de l’épisome n’est transférée que rarement puisque
sa deuxième moitié n’est transférée qu’en dernière position, si il n’y a pas eu
d’interruption, ce qui explique que les réceptrices ne deviennent jamais (en fait rarement)
F+.
Dans une coculture de souches F+ et F–, seul l’épisome est transféré, sauf chez
quelques donatrices devenues spontanément Hfr, ce qui explique, d’une part, qu’on
puisse observer des transferts de gènes alors que seul l’épisome est supposé pouvoir
être transféré, d’autre part, que ce transfert est beaucoup moins efficace puisque la
sous-population de donatrice Hfr est très petite parmi les bactéries F+.
Cela explique aussi pourquoi le transfert d’un gène particulier, à partir de donatrices
F+, ne donne jamais de résultats répétables puisque, d’une coculture à l’autre,
la sous-population de Hfr spontanées est hétérogène et que le gène particulier sera
plus ou moins efficacement transféré en fonction de l’éloignement du site d’insertion
de l’épisome et de son sens d’insertion dans les quelques Hfr spontanées, différentes
les unes des autres pour le site et le sens d’insertion.
En revanche, avec une souche « pure » Hfr, toutes les bactéries de la souche ont
leur épisome intégré au même site et dans la même orientation, ce qui conduit à des
résultats répétables lors du croisement avec une réceptrice puisque ce sont toujours
les mêmes gènes qui sont transférés dans le même ordre.
Jacob et Wollman ont tiré parti de ce fait pour établir une méthode de cartographie
des gènes bactériens par « conjugaison interrompue » selon le principe développé
dans l’exemple suivant.
On croise une coli Hfr sauvage, prototrophe pour l’arginine et la proline, et
sensible à la streptomycine, avec une réceptrice auxotrophe pour ces deux acides
aminés, et résistante à cet antibiotique.
Puis on prélève, toutes les minutes, deux
petits volumes de la coculture qu’on étale, après les avoir fortement agités afin de
rompre les ponts cytoplasmiques, le premier sur un milieu minimum additionné de
streptomycine et de proline, le deuxième sur un milieu minimum additionné de
streptomycine et d’arginine.
On observe le résultat suivant, des colonies apparaissent sur le premier milieu à
partir du quatrième prélèvement, tandis que des colonies n’apparaissent sur le
second milieu qu’à partir du dixième prélèvement.
L’interprétation suit alors :
– il faut attendre quatre minutes pour voir apparaître des recombinants [arg+], toute
conjugaison interrompue avant quatre minutes ne permet pas d’en avoir; le site
muté chez la réceptrice est donc localisé entre trois et quatre minutes de temps
minimal de conjugaison, à partir du site de l’origine de transfert de la Hfr;
– il faut attendre dix minutes pour voir apparaître des recombinants [pro+], toute
conjugaison interrompue avant dix minutes ne permet pas d’en avoir; le site muté
chez la réceptrice est donc localisé entre neuf et dix minutes de temps minimal de
conjugaison à partir du site de l’origine de transfert de la Hfr;
– la distance entre les sites de mutation arg et pro est égale à 6 minutes (environ
240 000 pb puisqu’il faut environ 100 minutes pour faire passer les 4,2 millions
de pb du génome de coli, soit environ 40 000 pb par minute).
Remarque. La streptomycine joue le rôle de marqueur de sélection des réceptrices
et permet de bloquer la croissance des Hfr sauvages prélevées dans la
coculture et qui, en absence de l’antibiotique, donneraient des colonies dans
toutes les boîtes d’étalement, les rendant ininterprétables.
C’est à partir de ce protocole simple que Jacob et Wollman ont cartographié les
centaines de mutations différentes d’incapacité de croissance sur lactose dans les
recherches qui les conduisirent à la définition de leur modèle de l’opéron lactose.
La parasexualité bactérienne conduit aussi à une conséquence importante et utile
pour l’analyse génétique, la sexduction.
Il arrive en effet qu’un souche Hfr redevienne spontanément F+ par excision de
son épisome.
Dans la très grande majorité des cas, cette excision est parfaite et
reconstitue le chromosome, d’une part, et l’épisome, d’autre part, mais dans de très
rares cas, du fait de l’homologie et de la dispersion des séquences d’insertion de
l’épisome, l’excision englobe à la fois la séquence de l’épisome et une séquence
adjacente du chromosome.
L’épisome est alors dénommé F′(x) ou x représente le
fragment de chromosome restant intégré à l’épisome.
Si une souche F′(x) conjugue avec une réceptrice ayant un chromosome entier, la
réceptrice va alors acquérir cet épisome et devenir « diploïde partiel » (on dit aussi mérodiploïde) pour la portion x du chromosome bactérien.
Cette diploïdie partielle
va permettre, pour les gènes localisés dans cette portion x, de réaliser les tests de
dominance ou de complémentation fonctionnelle qu’on aurait pu croire réservés à la
seule génétique eucaryote.
C’est par sexduction avec des épisomes F′(lac) que Jacob et Monod ont caractérisé
les mutants « récessifs ou dominants » de la région lactose qui leur permirent
d’établir le modèle de l’opéron lactose.
Remarque. Il est nécessaire de bien noter que les mérodiploïdes obtenus par
sexduction sont stables et formeront des clones de mérodiploïdes, tandis que
la conjugaison (ou, voir plus bas, la transduction et la transformation) ne
saurait conduire à des mérodiploïdes.
En effet, la réceptrice d’un épisome
F′(x) reçoit un réplicon capable de se répliquer de façon plus ou moins
synchrone avec le chromosome, tandis que la réceptrice d’un fragment
linéaire d’ADN (conjugaison, transduction ou transformation) ne reçoit pas
un réplicon, cet ADN exogène recombinera plus ou moins partiellement avec
l’ADN endogène ou disparaîtra, mais on n’obtiendra jamais un diploïde
partiel.