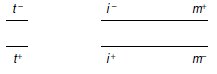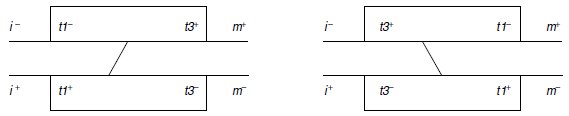1. Les diploïdes du croisement A × B donnent 50 % de spores [ilv+] et 50 % de spores [ilv–],
le mutant A diffère de B pour un seul des gènes impliqué dans la fraction commune des
chaînes de biosynthèse de l’isoleucine et de la valine. De même la ségrégation 2/2 pour le
phénotype méthionine permet de conclure que B est un mutant simple.
En revanche, les deux gènes mutés chez A et B sont génétiquement liés puisque la méiose
laisse apparaître des fréquences de spores recombinées sauvages ou double mutantes très
inférieures à celles des gamètes parentaux.
La fréquence des spores recombinées est égale à
260/9 000 = 0,0288; la distance entre les deux gènes (plus exactement les deux sites de
chacun des deux gènes) est égale à 2,88 ur.
2. Les quatre mutants ont un phénotype [trp–] récessif, ce qui permet de conclure qu’il n’y a
pas de complémentation fonctionnelle chez les diploïdes issus des croisements de mutants
entre eux, et que ces mutants sont mutés au moins dans un même gène (le même gène s’ils ne
sont mutés que dans un seul gène).
3. Toutes les spores peuvent pousser sur le milieu d’étalement, quel que soit leur phénotype,
mais seules les spores [trp+] peuvent pousser sur le milieu de réplique qui sert à tester la
ségrégation pour le seul phénotype tryptophane. Il y a ségrégation 2/2 pour les quatre
mutants qui sont donc mutés dans le même gène, mais sans doute pas au même site, puisque
ce sont des mutants indépendants.
4. Si le gène impliqué dans le phénotype tryptophane, noté t, était génétiquement indépendant
des deux gènes (très liés entre eux) impliqués dans les phénotypes ilv et met, notés i
et m, le génotype du diploïde pourrait s’écrire :
On devrait alors avoir égalité des phénotypes parentaux [ilv–, met+] et [ilv+, met–] parmi les
spores de phénotypes [trp+], ce qui n’est pas le cas (2 contre 4 836 !); le gène t est donc lié
aux deux autres et le génotype du diploïde peut s’écrire de trois façons différentes puisqu’on
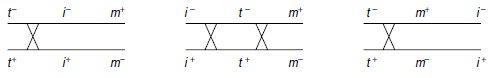
ne sait pas lequel des trois gènes est central (fig. 6.3).
Figure 6.3 bis Disposition des 3
gènes, selon trois ordres possibles.
Parmi les spores de phénotype [trp+], les spores [ilv–, met+] sont minoritaires, ce qui signifie
que l’ordre des gènes est tel que les spores les plus rares sont les spores de génotype (t+, i–, m+).
Si la première cartographie correspond à la réalité, les spores de génotype (t+, i–, m+), étant
issues d’un crossing-over entre les gènes t et i, seront plus fréquentes que les spores (t+, i–, m–)
nécessitant deux crossing-over.
Les observations étant non conformes aux résultats attendus
sous cette cartographie, celle-ci doit être rejetée.
Si la troisième cartographie correspond à la réalité, les spores de génotype (t+, i–, m+), étant
issues d’un crossing-over entre les gènes t et m, seront plus fréquentes que les spores (t+, i+, m+)
nécessitant deux crossing-over.
Les observations étant non conformes aux résultats attendus
sous cette cartographie, celle-ci doit être rejetée.
Si la deuxième cartographie correspond à la réalité, les spores de génotype (t+, i–, m+), étant
issues de deux crossing-over, entre les gènes t et i, et entre les gènes t et m, seront plus rares
que tous les autres types de spores recombinées ne nécessitant qu’un seul crossing-over.
Les
observations étant conformes aux résultats attendus sous cette cartographie, celle-ci peut être
acceptée, le gène t est central.
5. Chaque diploïde est porteur de deux exemplaires mutés du gène t, schématisé par un
rectangle, mais à des sites différents.
Le diploïde pourra être schématisé de deux façons
possibles (fig. 6.4), selon la disposition respective des sites t1– et t3– par rapport aux deux
gènes i et m.
Figure 6.4 Disposition relative des sites mutés du gène t par rapport aux gènes i et m.
Les notations t1–, t1+, t3– et t3+ se rapportent à la nature mutée ou sauvage de la
séquence nucléotidique en ce site précis du gène, celui-ci étant de toute façon non
fonctionnel quel que soit le site muté. On a figuré l’échange chromatidique permettant
de reconstituer une séquence sauvage fonctionnelle pour le gène t.
La figure 6.4 ne laisse figurer que l’échange chromatidique conduisant à une spore de phénotype
[trp+], ce qui permet bien de voir que, selon la disposition des sites t1 et t3, cette spore sauvage sera le plus souvent, à moins d’un autre crossing-over, possible mais rare, [ilv+, met+]
si la cartographie correspond à la première hypothèse, et [ilv–, met–] dans l’autre cas.
Des données du tableau 6.4, on peut conclure que les sites t1 et t3 correspondent au premier
type de cartographie.
L’ordre est ainsi i-t1-t3-m.
Pour t1 et t4, on obtient, l’ordre i-t4-t1-m,
d’où on peut conclure à l’ordonnancement i-t4-t1-t3-m. L’analyse des autres données permet
de conclure à la cartographie i-[t2, t4, t1, t3]-m.
Exercice 6.4
Des études génétiques ont montré que trois des gènes de levure, impliqués
dans la métabolisation du galactose sont contigus. Ils sont respectivement
nommés GAL7, GAL10 (central) et GAL1.
On a isolé, par irradiations aux rayons X, trois mutants [gal–], nommés d1,
d2 et d3, dont on peut suspecter qu’ils sont porteurs d’une délétion, d’une
part en raison du mutagène utilisé, mais aussi parce qu’ils ne donnent
aucun révertant, enfin parce que le diploïde issu du croisement
avec sauvage donne une stricte ségrégation 2/2 (50 % de spores [gal+] et
50 % de spores [gal–]) alors que ces mutants appartiennent simultanément
à deux groupes de complémentation entre lesquels il est assez facile
d’avoir des recombinaisons pour les mutants simples.
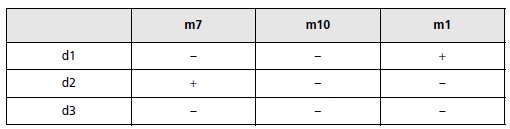
1. On croise les mutants d avec des mutants ponctuels simples touchés
respectivement dans l’un des trois gènes et désignés par m7, m10 et m1, et
on teste la capacité des diploïdes de pousser sur galactose. Interprétez les
résultats (tabl. 6.5).
TABLEAU 6.5 PHÉNOTYPE GAL DES DIPLOÏDES ISSUS DES CROISEMENTS
ENTRE MUTANTS PONCTUELS ET MUTANTS PAR DÉLÉTION.
« + » désigne la capacité de croissance sur galactose.
2. Quatre mutants ponctuels de GAL7, nommés m7-1, m7-2, m7-3 et m7-4
sont croisés soit avec le mutant d1, soit avec d3.
On met les diploïdes ainsi
obtenus à sporuler afin de recueillir un très grand nombre de spores qu’on
étale sur un milieu ne contenant que du galactose comme source de
carbone. Interprétez les résultats (tabl. 6.6).
Par ailleurs, les diploïdes issus des croisements entre le mutant ponctuel
m10-5 et le mutant d1 ou le mutant d2 sont capables de donner des
spores [gal+]. Concluez.
TABLEAU 6.6 CAPACITÉ DE CROISSANCE SUR GALACTOSE (NOTÉE +) DE SPORES ISSUES PAR MÉIOSE
DE DIPLOÏDES FORMÉS PAR CROISEMENTS ENTRE MUTANTS PONCTUELS DU GÈNE GAL7
ET MUTANTS PAR DÉLÉTION D1 OU D3.
– Cartographie par délétion pour ordonner des sites ponctuels et préciser l’amplitude
des délétions.
– Distinguer le test fonctionnel et le test de recombinaison.
Solution
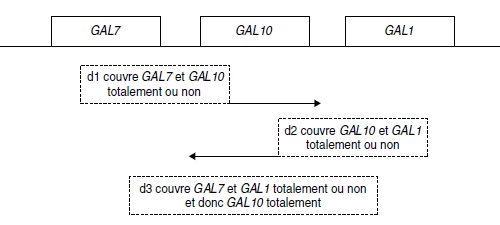
1. Il s’agit ici, par un test fonctionnel, de dénombrer les gènes « couverts » par la délétion;
l’analyse du tableau conduit au schéma suivant, où les cadres indiquent les étendues minimales
des délétions et les flèches les limites d’étendue des délétions, lorsqu’elles sont
identifiables.
2. Selon que la délétion portée par le diploïde ne couvre pas ou couvre le site de mutation
ponctuelle porté par la séquence allélique du gène étudié, on aura ou on n’aura pas de recombinant
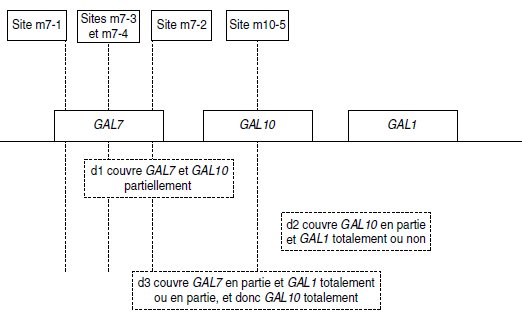
[gal+], ce qui permet de conclure, d’après le tableau 6.6, que les sites m7-2, m7-3
et m7-4 sont couverts par d1 alors que d3 ne couvre que m7-2, de sorte qu’on peut déduire
l’ordre des sites (sauf pour m7-3 et m7-4) et préciser que d3 n’est pas aussi étendue dans
GAL7 que d1.
La capacité de production de spores [gal+] dans les croisements entre m10-5 et d1 ou d2
prouve que ce site n’est couvert par aucune des deux délétions qui ne sont donc pas
chevauchantes.