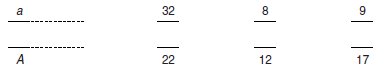Ordonner des gènes ou des sites sur un axe peut
être réalisé par le calcul de leurs distances respectives, ce
qui suppose que le nombre de gamètes étudiés est d’autant plus
élevé que les distances sont courtes.
Si les conditions
expérimentales limitent ce nombre et ne permettent pas d’estimer
avec précision les distances, il peut être utile de définir un
test, plus qualitatif que quantitatif, fondé sur l’observation
d’un type particulier de gamètes dans des croisements
parallèles, les souches croisées étant définies de manière telle
que ce gamète sera très rarement, voire jamais formé dans un des
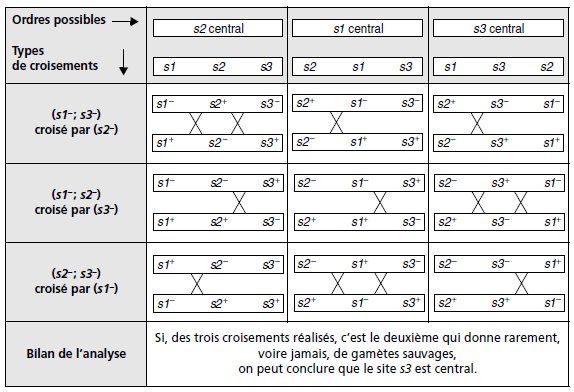
croisements. Supposons que trois sites de mutations d’un même
gène, notés s1, s2 et s3 doivent être ordonnés, trois ordres
sont possibles selon que s1 ou s2 ou s3 est central.
Dans un tel
but on peut, à condition d’en disposer, faire trois croisements
parallèles entre un double mutant pour deux des sites et le
simple mutant pour le troisième, et observer lequel des
croisements ne donne jamais de gamètes sauvages; on peut alors
en conclure que le site du mutant simple de ce croisement est le
site central (tabl. 6.1).
Ce genre de
croisement et de test des gamètes est assez facile chez la
levure où des milliers de spores peuvent être déposées sur une
boîte où seules les sauvages peuvent donner des colonies, ou
chez la drosophile si un test cross permet aux seules sauvages
F2 de se développer.
Il existe
des variantes plus simples, notamment quand on ordonne deux à
deux les sites d’un même gène par rapport à une mutation externe
responsable d’un phénotype différent, pouvant lui-même servir de
crible de sélection (voir exercice d’application et de génétique
bactérienne); dans ce cas il y a deux cartes possibles (le
marqueur externe ne pouvant être central) et il convient de
comparer les résultats de deux croisements.
TABLEAU 6.1 TEST TROIS POINT
PERMETTANT D’ORDONNER LES MARQUEURS. Le but est d’identifier, en
fonction de l’ordre possible des 3 marqueurs, le croisement
entre doubles et simples mutants où la formation d’un gamète
sauvage est la plus rare, parce qu’exigeant un double
crossing-over.
On peut
enfin définir un test quatre points où on ordonne deux à deux
des sites de mutations en les positionnant par rapport à deux
mutations extérieures situées de part et d’autre (voir
exercices).
La plupart
du temps, ces tests nécessitent de comparer entre eux les
résultats de plusieurs croisements parallèles, ce qui n’a de
sens que si les observations ne dépendent que de la position des
sites et d’aucun autre paramètre; on verra que dans certaines
conditions, avec des marqueurs de sélection supplémentaires, on
peut raisonner à l’intérieur d’un seul croisement et
s’affranchir ainsi des paramètres qui, en dehors de la position
des sites, sont susceptibles de jouer sur l’efficacité du
croisement à générer les recombinants (voir exercice de
génétique bactérienne).
Exercices :
Exercice 6.1
En 1986, le
gène humain impliqué dans la mucoviscidose, appelé depuis CFTR
(Cystic Fibrosis Transmembrane conductance Regulator) n’avait
pas encore été cloné et sa fonction était encore inconnue, même
si elle était suspectée (canal ionique chlorure).
Par analyse
de liaison génétique, on a montré que le gène CFTR était lié
(distance égale à 15 cM) à un marqueur polymorphe de l’ADN dont
les fragments de digestion (par l’enzyme Hinc II) sont reconnus
par une sonde spécifique LAM4-917, mais ce marqueur polymorphe,
nommé DOCRI- 917 n’était pas encore lui-même assigné à un
chromosome.
On a alors
extrait l’ADN de plusieurs lignées hybrides homme-rongeur qu’on
a digéré par Hinc II; les fragments ont été ensuite séparés par
électrophorèse puis transférés, après dénaturation de l’ADN, sur
une membrane de nylon; ces Southern blot ont été hybridés par la
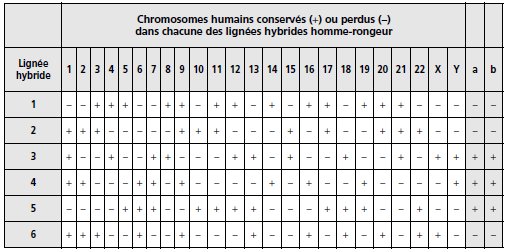
sonde marquée LAM4-917 (tabl. 6.2, colonne a).
Par la
suite, on a réhybridé les Southern blot par une autre sonde
marquée, TCRB, correspondant à la séquence du gène du récepteur
β de lymphocyte T (tabl. 6.2, colonne b), localisé sur le
chromosome 7. Interprétez ces résultats en justifiant les choix
de sondes.
TABLEAU 6.2 ASSIGNATION
CHROMOSOMIQUE DU GÈNE CFTR. Colonne a, signal d’hybridation de
l’ADN des lignées avec la sonde LAM4-917; colonne b, signal
d’hybridation avec la sonde TCRB. (+) indique un signal
d’hybridation et (–) son absence.
Solution. Le
but de ce protocole est d’assigner le gène CFTR à un chromosome
en assignant le marqueur DOCRI-917 qui lui est génétiquement
donc physiquement lié.
Le gène CFTR
ne peut être directement utilisé pour cette localisation,
puisque sa séquence est indisponible, le gène n’étant pas encore
cloné, et que son produit encore inconnu ne peut être dosé dans
des extraits acellulaires de cellules hybrides.
On attend,
de l’hybridation avec la sonde, un signal positif si le
chromosome porteur de DOCRI-917 est présent, et l’absence de
signal s’il est absent.
L’incohérence de résultat pour un chromosome donné (signal
d’hybridation présent en absence de ce chromosome chez l’hybride
cellulaire, ou signal d’hybridation absent en présence de ce
chromosome) exclut alors ce chromosome comme porteur du marqueur
DOCRI-917 et donc du gène CFTR.
La sonde
LAM4-917 donne un signal d’hybridation avec l’ADN des lignées 3,
4 et 5 et un signal négatif avec les autres.
Si on
considère les chromosomes conservés ou perdus par les
différentes lignées, les résultats observés sont toujours
incohérents, sauf avec le chromosome 7, ce qui permet de
localiser DOCRI-917 et le gène CFTR sur le chromosome 7.
L’hybridation avec le gène TCRB joue le rôle de contrôle,
montrant que les Southern blot répondent bien de façon attendue,
positive ou négative, à l’hybridation d’une séquence connue pour
être localisée sur le chromosome 7.
Exercice 6.2
On a isolé
un mutant albinos dans une lignée pure de souris nommée CD1.
Une étude
génétique a permis de montrer que ce mutant était récessif et
muté dans un seul gène (3/4 de sauvage et 1/4 d’albinos à
l’issue d’un croisement F1 × F1).
On souhaite
« assigner » ce gène à un chromosome, c’est-à-dire identifier le
chromosome où réside le locus de ce gène.
Dans ce but,
on croise des mutants albinos de la lignée CD1 avec des
individus de la lignée 129, sachant que ces deux lignées
diffèrent l’une de l’autre pour de nombreux marqueurs VNTR
identifiés et cartographiés.
Les
marqueurs VNTR (Variable Number of Tandem Repeats) sont des
séquences d’ADN formées d’un nombre variable d’un motif répété,
souvent un di ou un trinucléotide.
Ce «
polyallélisme » génère dans une population naturelle un grand
nombre de génotypes différents les uns des autres et entre eux,
ce qui constitue la base des méthodes d’empreintes génétiques.
Dans les
lignées pures de souris, tous les individus sont homozygotes
pour un allèle du marqueur mais les lignées diffèrent les unes
des autres, les individus n’étant pas homozygotes pour le même
allèle du marqueur.
Les F1 sont
croisées entre elles et on récupère les F2 albinos; on
entreprend alors une « revue génomique » (génome scan) qui
consiste à déterminer, pour tous ces individus, le génotype dont
ils sont porteurs pour toute une série de ces marqueurs
moléculaires répartis sur les différents chromosomes, dont les
marqueurs D4M24, D5M8 et D7M52 (tabl. 6.3).
– D4M24 est
le marqueur 24 du chromosome 4, les individus CD1 étant
homozygotes pour l’allèle porteur de 32 répétitions, les
individus 129 étant homozygotes pour l’allèle porteur de 22
répétitions;
– D5M8 est
le marqueur 8 du chromosome 5, les individus CD1 étant
homozygotes pour l’allèle porteur de 8 répétitions, les
individus 129 étant homozygotes pour l’allèle porteur de 12
répétitions;
– D7M52 est
le marqueur 52 du chromosome 7, les individus CD1 étant
homozygotes pour l’allèle porteur de 9 répétitions, les
individus 129 étant homozygotes pour l’allèle porteur de 17
répétitions.
TABLEAU 6.3
GÉNOTYPES DES TROIS MARQUEURS VNTR ÉTUDIÉS DES F2 DE PHÉNOTYPE
ALBINOS. Les allèles de chacun des marqueurs sont définis par
leur nombre de répétitions, (entre parenthèses, effectifs
observés de chacun des génotypes).
Sur quel
chromosome peut-on assigner la mutation albinos de la lignée CD1
? Justifier les réponses par un schéma.
Solution. La
méthode consiste à assigner un gène à un chromosome en montrant
qu’il est génétiquement lié à un marqueur moléculaire connu de
ce chromosome. Les croisements entre albinos CD1 et non-albinos
129 génère des hétérozygotes pour tous les gènes, et notamment
les marqueurs, dont l’allèle est différent d’une lignée pure à
l’autre; c’est le cas pour le gène impliqué dans l’albinisme,
dont les allèles seront notés A et a, comme pour les trois
marqueurs étudiés.
Le génotype
des F1 est figuré ci-dessous; les pointillés désignent une
éventuelle liaison avec l’un des trois marqueurs (les trois
étant physiquement indépendants entre eux) :
En cas
d’indépendance génétique entre le couple d’allèles A/a et un
marqueur donné, les F2, qu’ils soient A//A, A//a ou a//a seront,
pour le marqueur considéré, homozygotes pour l’un des allèles
avec une probabilité égale à 1/4 et hétérozygotes avec une
probabilité égale à 1/2.
C’est
effectivement ce qu’on observe pour D4M24 et D7M52; attention
cette observation, en elle-même ne permet pas d’exclure que le
gène A soit sur le chromosome 4 ou sur l7, mais permet d’exclure
qu’il soit sur le 4, dans le voisinage de D4M24 et sur le 7,
dans le voisinage de D7M52.
N’oublions
jamais que deux gènes ou marqueurs peuvent être physiquement
liés tout en étant génétiquement indépendants.
En cas de
liaison génétique entre le couple d’allèles A/a et un marqueur
donné, les allèles A auront tendance à coségréger, à la méiose
chez la F1, avec l’allèle marqueur du parent 129, et les allèles
a auront tendance à coségréger avec l’allèle marqueur de la
lignée CD1.
En
conséquence, les albinos F2, de génotype a//a, seront beaucoup
plus souvent homozygotes pour l’allèle CD1, parfois
hétérozygotes, et plus rarement homozygotes pour l’allèle 129,
puisqu’alors ils seraient issus de deux gamètes, paternel et
maternel, résultant tous deux d’un crossing-over entre le gène
et le marqueur.
C’est ce
qu’on observe pour le marqueur D5M8; on peut en conclure que la
mutation albinos de CD1 touche un gène du chromosome 5, dans le
voisinage du marqueur D5M8.
Exercice 6.3
Dans tout
l’exercice, on ne tiendra pas compte du type sexuel, a ou α, des
souches de levure Saccharomyces cerevisiae, on suppose qu’on
dispose toujours d’une souche du type sexuel requis pour le
croisement, ainsi que des marqueurs de sélection des diploïdes.
On dispose
d’une souche haploïde A, auxotrophe pour l’isoleucine et la
valine, phénotype noté [ilv–] deux acides aminés dont la chaîne
de biosynthèse comprend une partie commune, et d’une souche B,
auxotrophe pour la méthionine, phénotype noté [met–].
1. On
réalise le croisement de A par B puis on étudie les spores
issues de la méiose des diploïdes; on observe 4 250 spores
[ilv–], 4 230 spores [met–], 140 spores [ilv–, met–] et 120
spores [ilv+, met+]. Quelles conclusions peut-on en tirer ?
2. À partir
de la souche A, on a isolé un grand nombre de mutants
indépendants, auxotrophes pour le tryptophane, phénotype noté
[trp–]; on étudie quatre mutants nommés t1, t2, t3 et t4.
– Les quatre
mutants sont croisés avec la souche B, les diploïdes sont
sauvages.
– Les quatre
mutants sont croisés deux à deux, les diploïdes sont tous
[trp–].
Que conclure
?
3. Les
diploïdes issus du croisement du mutant t1 avec B sont mis à
sporuler.
On étale
environ 10 000 spores sur des boîtes de milieu Mo additionné de
tryptophane, de méthionine, de valine et d’isoleucine; 4 980
colonies sont capables de pousser, après réplique sur Mo
additionné de méthionine, de valine et d’isoleucine.
On obtient
des résultats sans différence significative avec les autres
mutants t2, t3 et t4. Concluez.
4. Parmi ces
4 980 colonies, 4 836 se révèlent [ilv+, met–], 69 sont [ilv+,
met+], 73 sont [ilv–, met–] et 2 sont [ilv–, met+]. Donnez la
disposition des gènes entre eux sans faire de calculs mais en
détaillant le génotype du diploïde dont sont issues les spores
étudiées.
5. À partir
des croisements précédents t3 × B et t4 × B, on a pu isoler des
spores de phénotype [met–, trp–, ilv+] qui sont respectivement
nommées t3′ et t4′. Précisez leur génotype.
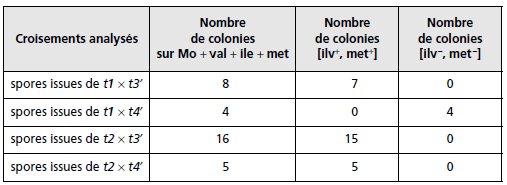
On croise
chaque mutant t1 et t2 avec chaque mutant t3′ et t4′; les
diploïdes sont mis à sporuler et on étale environ 10 000 spores
issues de chacun des croisements sur des boîtes de milieu Mo
additionné de méthionine, de valine et d’isoleucine.
Par réplique
on teste les colonies alors obtenues pour les phénotypes [ilv]
ou [met]. Interprétez les résultats (tabl. 6.4).
TABLEAU 6.4 EFFECTIFS DES
COLONIES CAPABLES DE POUSSER SUR CHACUN DES MILIEUX DE RÉPLIQUE
À PARTIR DE BOÎTES MÈRES CONTENANT 10 000 COLONIES ISSUES DES
SPORES OBTENUES À PARTIR DES QUATRE CROISEMENTS ANALYSÉS.
–
Cartographie de gènes chez la levure Saccharomyces cerevisiae
par test trois points.