•
Les cellules
cancéreuses ont 2
caractéristiques :
elles se divisent
sans être soumises
aux contrôles
normaux de la
croissance
cellulaire et sont
capables d’envahir
des tissus
normalement réservés
à d’autres cellules.
• La
transformation
cancéreuse résulte
de lésions
génétiques multiples
sur des gènes
régulant la
croissance et la
différenciation
cellulaire : les
proto-oncogènes
normaux qui vont
subir des mutations
activatrices dans
les cellules
cancéreuses, les
transformant ainsi
en oncogènes
(capables notamment
d’entraîner une
division cellulaire
inappropriée) et les
anti-oncogènes, ou
plutôt gènes
suppresseurs de
tumeurs, détruits ou
non fonctionnels
dans les cellules
cancéreuses. Le
cancer est
essentiellement une
maladie des gènes.
• La
plupart des
oncogènes codent
pour des protéines
impliquées dans la
transmission du
signal mitotique
induit par les
facteurs de
croissance de la
membrane jusqu’au
noyau. Ces facteurs
fréquemment produits
en excès par les
cellules cancéreuses
vont parfois jouer
des rôles multiples
: facteur de survie
(antiapoptotique),
facteur mitogénique,
de motilité ou
angiogénique.
Les lésions des oncogènes et des gènes suppresseurs de tumeur affectent l’interaction des cellules tumorales avec leur environnement normal et permettent leur dissémination dans l’organisme, en altérant la structure ou la fonction des molécules d’adhésion cellulaire, en induisant la production de facteurs de motilité et d’enzymes capables de détruire la matrice extracellulaire. Ces altérations permettent aux cellules tumorales de coloniser et de se diviser dans les tissus où elles ne résident pas normalement.
Phénotype et clonalité des
cellules cancéreuses :
Quel que soit leur tissu
d’origine, les cellules cancéreuses présentent des anomalies phénotypiques et
fonctionnelles communes.
La taille de la cellule
cancéreuse et de son noyau est fréquemment augmentée par rapport à la cellule
normale et varie au sein de la population cellulaire tumorale (anisocytose,
anisocaryose).
Elle conserve souvent une
morphologie arrondie comme les cellules en mitose. Son noyau présente une
chromatine irrégulière et la proportion des cellules en mitose (index mitotique)
est augmentée.
Sur le plan fonctionnel,
les cellules cancéreuses présentent des altérations de l’expression ou de la
fonction de molécules d’adhésion à la matrice extracellulaire, une
réorganisation du cytosquelette d’actine du cortex cellulaire, une motilité
accrue, une augmentation de la production de facteurs de croissance,
d’activateurs du plasminogène et d’enzymes capables de dégrader les composants
de la matrice.
Les principales anomalies
fonctionnelles des cellules cancéreuses portent cependant sur leurs capacités de
croissance et de division : elles peuvent se diviser en présence de très faibles
concentrations de facteurs de croissance et ont perdu la capacité d’arrêter de
se diviser lorsque la densité cellulaire est élevée et que les cellules sont en
contact les unes avec les autres (perte de l’inhibition de contact).
Elles sont capables de se
diviser sans attache avec une surface solide (perte de la dépendance d’ancrage)
et peuvent pousser dans des milieux semisolides tels que l’agar mou.
Enfin, elles sont capables
d’un nombre illimité de divisions cellulaires sans subir le phénomène de
sénescence (immortalisation) et d’induire des tumeurs après injection chez des
animaux susceptibles (souris athymiques).
Bien que le terme «
transformation » ne désigne stricto sensu que la modification transmissible à la
descendance des propriétés d’une cellule eucaryote, dans le cas des cellules
animales en culture, le terme « transformation » est fréquemment utilisé pour
désigner l’acquisition de propriétés analogues à celles des cellules
cancéreuses.
On classe les cancers
selon le tissu et le type cellulaire dont ils proviennent.
On distingue ainsi les
carcinomes (tumeurs malignes issues de cellules épithéliales) qui sont les
cancers les plus fréquents de l’adulte ; les sarcomes (cancers provenant des
cellules du tissu conjonctif, spécialisé ou non) ; les tumeurs du système
hématopoïétique (leucémies ou lymphomes) ; les tumeurs du système nerveux
central ; les tumeurs embryonnaires… Dans ces différents types cellulaires, les
voies de contrôle de la différenciation et de la croissance cellulaire utilisent
souvent des mécanismes moléculaires communs.
La cancérisation de la
cellule est la conséquence de mutations de gènes codant pour des protéines
impliquées dans ces différentes voies de régulation.
Les cancers correspondent
le plus souvent à la prolifération d’un clone cellulaire dérivant d’une seule
cellule anormale.
Cela a pu être démontré
chez les femmes par l’analyse de l’origine paternelle ou maternelle du
chromosome X inactivé dans les cellules tumorales qui montre que toutes les
cellules tumorales ont le même chromosome X inactivé.
Cela a pu être confirmé
plus finement par l’analyse de la séquence nucléotidique des sites de
translocations chromosomiques spécifiques, telles que la t(9,22) dans la
leucémie myéloïde chronique (LMC), qui sont toujours identiques dans une même
tumeur chez un individu, mais le plus souvent différentes dans deux tumeurs de
même histologie chez deux individus différents.
Ces observations
démontrent l’origine clonale de la plupart des cancers.
Il existe cependant
quelques exceptions et certaines proliférations cellulaires tumorales, notamment
des lymphomes de sujets immunodéprimés, peuvent comporter une population
cellulaire polyclonale ou oligoclonale.
Anomalies génétiques des
cellules cancéreuses :
MUTATIONS DE L'ADN
NÉCESSAIRES POUR LA TRANSFORMATION CANCÉREUSE :
Des mutations de l’ADN de
la cellule sont à l’origine de la majorité des cancers. Il existe une
corrélation entre le potentiel mutagène pour l’ADN de produits chimiques
(méthylcholanthrène, 2-naphtylamine) et leur potentiel carcinogène. Il est
cependant rare qu’une seule mutation soit suffisante pour entraîner la
transformation maligne.
En général, plusieurs
mutations dans des gènes différents sont requises pour l’acquisition d’un
phénotype cancéreux. On estime ainsi qu’il faut entre 3 et 7 mutations
indépendantes pour transformer une cellule normale en cellule cancéreuse.
La nécessité de plusieurs
mutations indépendantes pour la transformation d’une cellule explique le délai
souvent long (plusieurs années) entre l’exposition à un carcinogène et
l’apparition d’un cancer.
Au cours de cet
intervalle, les cellules initialement mutées acquièrent des mutations
supplémentaires dans d’autres gènes qui contribuent à la transformation
cancéreuse.
Chez les individus non
exposés à des carcinogènes, la transformation maligne survient au hasard et
provient de plusieurs mutations aléatoires dans une même cellule, qui mettent
des années à se constituer.
De ce fait, l’incidence de
la plupart des cancers augmente de manière exponentielle avec l’âge.
Chez les individus
porteurs d’une mutation constitutionnelle de certains gènes, notamment de gènes
suppresseurs de tumeurs tels que p53, APC ou BRCA-1 (v. infra), les cancers
surviennent plus précocement, une première « étape » dans le processus de
transformation cancéreuse ayant été franchie dans toutes les cellules somatiques
de l’individu.
Le développement d’un
cancer peut également être favorisé par des facteurs qui n’induisent pas de
mutation de l’ADN mais stimulent la prolifération cellulaire ou bloquent la
différenciation.
Ces produits non mutagènes
sont appelés promoteurs tumoraux. Dans un premier temps, l’exposition a un
carcinogène induit une première lésion génétique latente. Cette étape est
parfois appelée initiation tumorale.
La lésion génétique ainsi
induite permet la transformation cancéreuse à la faveur soit de l’acquisition
d’anomalies génétiques supplémentaires telles que celles décrites ci-dessus,
soit après exposition répétée à ces promoteurs tumoraux qui vont stimuler la
division cellulaire ou bloquer le processus normal de différenciation. Comme
exemple de promoteurs tumoraux, on peut citer les esters de phorbol qui agissent
en activant la protéine kinase C.
Les corrélations
épidémiologiques entre les antécédents gynéco obstétricaux d’une patiente (âge
des premières règles, de la première grossesse, de la ménopause) et le risque
relatif de cancer du sein suggèrent que les hormones sexuelles, notamment les
œstrogènes, jouent un rôle de promoteur tumoral pour certains cancers du sein.
Au total, le processus de
transformation cancéreuse est la conséquence de l’accumulation de mutations de
l’ADN pour la plupart des tumeurs humaines et animales.
GENES IMPLIQUÉS DANS
LA TRANSFORMATION MALIGNE :
La progression tumorale
correspond à un phénomène dynamique qui, à chaque étape, sélectionne une cellule
ayant acquis un avantage de croissance (soit une activation de la multiplication
cellulaire, soit une inhibition de la mort cellulaire). Notre organisme est
composé d’environ 5 x 1012 cellules réparties dans plus de 200 types cellulaires
différents.
Leur multiplication et
leur durée de vie sont rigoureusement contrôlées : certaines cellules, telles
les cellules nerveuses, ne nécessitent pas un renouvellement constant, d’autres,
telles les cellules sanguines, sont perpétuellement en cours de division.
Le contrôle de la
croissance cellulaire résulte d’un équilibre permanent entre facteurs
stimulateurs et facteurs inhibiteurs de la croissance cellulaire. Au cours de la
vie cellulaire, l’ADN est soumis à des agressions qui peuvent résulter d’erreurs
de la machinerie cellulaire de réplication ou d’agents extérieurs comme les
agents génotoxiques carcinogènes mentionnés ci-dessus.
De façon générale, ces
altérations de l’ADN sont immédiatement réparées par des mécanismes spécialisés.
Cependant, dans de rares cas, la réparation ne se fait pas ou mal, et une
mutation génique apparaît.
Si cette mutation atteint
et modifie la structure d’un gène codant pour un facteur qui contrôle la
prolifération cellulaire ou le phénomène de mort cellulaire (apoptose), la
cellule peut acquérir un avantage de croissance à l’origine d’une expansion
clonale.
La description de gènes
impliqués dans la transformation maligne a permis d’élaborer une théorie unifiée
concernant les mécanismes moléculaires mis en jeu dans la genèse d’une tumeur.
Deux grandes catégories de
gènes peuvent ainsi être distinguées : les gènes dont les produits contribuent à
stimuler la prolifération cellulaire et ceux dont le produit réprime la
prolifération cellulaire.
Les premiers gènes sont
appelés en général oncogènes et les seconds anti-oncogènes ou plutôt gènes
suppresseurs de tumeur.
1. Oncogènes :
Si l’on exclut certains
oncogènes viraux (notamment de virus à ADN), les oncogènes sont le plus souvent
des gènes cellulaires mutés. Le terme proto-oncogène désigne le gène cellulaire
normal, qui est muté en oncogène dans une cellule tumorale, favorisant ainsi la
transformation cancéreuse de la cellule.
La mutation d’un oncogène
rend en général le gène « hyperactif » et ces mutations sont généralement
dominantes.
Les oncogènes ont été
découverts initialement dans des rétrovirus responsables de tumeurs animales. Le
premier rétrovirus ainsi individualisé est un virus du poulet, le virus de
sarcome de Rous.
Il contient un gène appelé
v-src, qui n’est pas indispensable à la réplication virale mais qui a été
prélevé accidentellement par le virus dans une cellule hôte antérieure. Le gène
viral v-src possède ainsi un homologue cellulaire normal, désigné c-src et qui
est présent dans le génome de la plupart des vertébrés. Le gène cellulaire c-src
code pour une protéine membranaire qui intervient dans la transduction du signal
mitotique.
Les modifications
qualitatives du gène src chez le virus provoquent une activation anormale de la
protéine, expliquant ses capacités transformantes. On s’est secondairement rendu
compte que beaucoup de tumeurs humaines, non induites par un virus, présentaient
des mutations activatrices du proto-oncogène c-src qui le transforment en un
oncogène. D’autres oncogènes contenus dans des rétrovirus responsables de
sarcomes ou de leucémies chez les oiseaux ou les mammifères ont été
individualisés.
Ces oncogènes viraux
(v-onc) possèdent un homologue cellulaire (c-onc), qui est muté dans certaines
tumeurs humaines non liées à des rétrovirus.
Ces oncogènes sont très
souvent impliqués dans la transmission d’un signal de prolifération cellulaire
du milieu extracellulaire jusqu’au noyau : ce sont des gènes codant pour des
facteurs de croissance ou cytokines (v-sis, homologue de la chaîne B du platelet
derived growth factor, PDGF), des récepteurs de cytokines (v-erb-B1, v-kit,
v-fms), des tyrosine kinases du feuillet interne de la membrane plasmique
(v-src), intracytosoliques (v-fes), des sérine thréonine kinases (v-raf), des
protéines fixant le guanosine triphosphate (GTP) (H-ras), des facteurs
régulateurs de la transcription (v-fos, v-jun, v-rel), des récepteurs d’hormone
liposoluble (v-erbA).
Tous les oncogènes n’ont
cependant pas été identifiés dans des rétrovirus.
D’autres oncogènes, et
donc d’autres proto-oncogènes, ont été identifiés dans les tumeurs humaines par
transfection de lignées de fibroblastes immortalisées NIH 3T3 par de l’ADN
provenant de lignées tumorales.
Au total, près d’une
centaine d’oncogènes ont désormais été identifiés. Les mutations observées dans
les proto-oncogènes modifient la séquence des acides aminés de la protéine en
modifiant ses propriétés fonctionnelles, lui conférant une activité permanente
et non régulable par les processus normaux de contrôle.
Les rétrovirus peuvent
également activer un proto-oncogène en s’insérant à proximité de ses séquences
régulatrices, conduisant ainsi à sa surexpression : on parle alors de mutagenèse
insertionnelle.
En fait, différents
mécanismes d’activation d’un oncogène cellulaire peuvent être observés.
• La mutation ponctuelle a
pour exemple classique la mutation des codons 12, 13 ou 61 du gène H-ras, qui
empêche son produit protéique d’hydrolyser le GTP, lui conférant ainsi une
activité biologique permanente.
• Lors de la délétion
d’une partie de la séquence codante, le gène c-erbB1 code pour le récepteur de
l’epidermal growth factor (EGF) ; dans certaines tumeurs, on observe une
délétion de la partie extracellulaire capable de fixer EGF et des mutations dans
sa portion intracytoplasmique conférant à la protéine une activité tyrosine
kinase permanente.
• La translocation
chromosomique va réunir dans une protéine de fusion le produit d’un gène
activement transcrit et une partie des séquences codantes de l’oncogène,
conduisant à la synthèse d’une protéine hyperactive ; un exemple de ce type de
fusion est la protéine bcr-abl, produit de la translocation t(9,22) de la
leucémie myéloïde chronique.
• Parfois, la
translocation va simplement mettre l’oncogène sous la dépendance du promoteur
d’un gène activement transcrit, comme le gène des chaînes lourdes des
immunoglobulines pour la t(8,14) des lymphomes de Burkitt.
• L’amplification génique
est décrite pour les gènes N-myc dans le neuroblastome.
• L’insertion d’un élément
génétique mobile (tel qu’un rétrovirus) à proximité du proto-oncogène entraîne
l’activation de la transcription du gène. Le gène codant pour le fibroblast
growth factor 3 (FGF-3) est ainsi activé par le mouse mammary tumour virus chez
la souris conduisant à l’apparition de tumeurs mammaires. Les mutations des
oncogènes ne surviennent pas isolément dans les tumeurs et très souvent, la
mutation de plusieurs oncogènes est synergique pour l’acquisition d’un phénotype
cancéreux.
Ce phénomène est connu
sous le nom de coopération des oncogènes.
2. Anti-oncogènes ou
gènes suppresseurs de tumeurs :
Certaines mutations vont
inactiver des gènes dont le produit est impliqué dans le blocage de l’entrée
dans le cycle cellulaire : on parle parfois à leur propos d’antioncogènes, mais
le terme gène suppresseur de tumeur est généralement préféré. Beaucoup de gènes
suppresseurs de tumeurs ont été identifiés grâce à l’étude des syndromes de
prédisposition héréditaire aux cancers.
C’est le cas du gène du
rétinoblastome (Rb), premier gène suppresseur de tumeur identifié (1984) grâce à
l’étude des familles atteintes de formes héréditaires de rétinoblastome. Dans la
forme héréditaire de cette maladie, les individus atteints présentent souvent
plusieurs tumeurs tandis que les formes sporadiques de rétinoblastome sont en
général des tumeurs uniques.
Le gène Rb a pu être
identifié dans une région du chromosome 13 touchée par une délétion chez les
individus atteints dans certaines de ces familles.
On a pu montrer que dans
les cellules de rétinoblastome, les 2 copies du gène sont inactivées, une 2e
mutation ayant inactivé le gène Rb sur l’autre chromosome dans la cellule
tumorale.
La probabilité de survenue
de cette 2e mutation est donc élevée dans ces cellules présentant toutes une
mutation du 1er gène Rb, et plusieurs tumeurs peuvent ainsi survenir dans les
cellules rétiniennes des 2 yeux ; en revanche chez les individus dépourvus de
mutation de Rb constitutionnelle, le risque de survenue d’une mutation sur
chacun des 2 gènes Rb dans la même cellule est très faible, expliquant ainsi la
rareté de cette affection en dehors des formes familiales.
La perte du gène Rb joue
un rôle important dans des tumeurs plus fréquentes que le rétinoblastome. Les 2
copies du gène sont fréquemment altérées dans les cancers du poumon, du sein, de
la vessie, les sarcomes.
La protéine Rb,
lorsqu’elle est déphosphorylée, inhibe la transcription de certains gènes,
permettant l’entrée dans le cycle cellulaire.
Lorsque les 2 copies du
gène sont perdues ou lorsque la protéine Rb est phosphorylée (v. infra), il
n’empêche plus la transcription et permet ainsi à la cellule d’accomplir une
nouvelle division.
Depuis la description
initiale du gène Rb comme modèle de gène suppresseur de tumeur, de nombreux
autres gènes suppresseurs de tumeurs ont été identifiés dans les tumeurs
humaines : APC et DCC dans les tumeurs coliques, WT1 dans les tumeurs de Wilms,
BRCA-1 et 2 dans les adénocarcinomes du sein, p53 dans une large variété de
cancers. La protéine p53 est mutée dans une large variété de cancers humains.
Une des fonctions de p53
est d’induire la production d’une protéine p21/WAF1 inhibitrice des kinases
dépendantes des cyclines qui permettent à la cellule de passer le point de
contrôle G1 du cycle cellulaire ; p53 est induite en réponse à des altérations
de l’ADN et permet à la cellule d’arrêter sa progression dans le cycle, de
réparer les lésions de l’ADN ou, si celles-ci sont irréparables, d’entraîner
l’apoptose de la cellule altérée.
Les mutations de p53
permettent à la cellule à la fois de se diviser et d’acquérir des mutations
supplémentaires sur d’autres gènes.
Les mutations
constitutionnelles de p53 sont responsables d’un syndrome de prédisposition
génétique aux cancers appelé syndrome de Li-Fraumeni. D’autres gènes
suppresseurs de tumeurs sont mutés chez des sujets porteurs de prédispositions
héréditaires au cancer.
La polypose colique
adénomateuse familiale résulte d’une mutation somatique d’un gène appelé APC ;
les gènes de réparation msh-2 et mlh-1 sont altérés dans les cancers coliques
familiaux non polyposiques, le gène WT1 dans les tumeurs de Wilms familiales.
Certains gènes
suppresseurs de tumeurs, comme p53 ou Rb, sont inactivés par d’autres mécanismes
que des mutations inactivatrices. Les virus oncogènes à ADN, tels que les
papillomavirus (impliqués dans les cancers du col de l’utérus) ou le virus
polyome contiennent des protéines virales (protéines E6 et E7 de papillomavirus,
antigène T et t de polyome) qui se fixent aux protéines p53 et à Rb bloquant
ainsi leur fonction.
Le virus peut ainsi
utiliser à son profit les protéines cellulaires de réplication de l’ADN de
l’hôte pour se répliquer. Dans certaines cellules, ces virus oncogènes à ADN
vont s’in¬tégrer à l’ADN cellulaire et les protéines virales sont produites en
permanence, bloquant ainsi en permanence la fonction des protéines Rb et p53 et
favorisant la division cellulaire. À la différence des rétrovirus, ces protéines
oncogènes des virus à ADN n’ont pas d’équivalent cellulaire et sont
indispensables à la réplication virale.
3. Modèle du carcinome
colique :
Plusieurs mutations de
proto-oncogènes ou de gènes suppresseurs de tumeurs sont donc requises pour la
transformation d’une cellule normale en cellule cancéreuse. La genèse des
anomalies moléculaires, de l’épithélium normal, à l’adénome puis à
l’adénocarcinome a été particulièrement bien étudiée dans les cancers
recto-coliques.
La plupart des cellules
d’adénocarcinome colique acquièrent au cours de leur transformation cancéreuse
des mutations sur des gènes précis : le gène suppresseur de tumeur APC est perdu
souvent avant l’apparition de l’adénome, l’oncogène K-ras est muté et le gène
suppresseur de tumeur DCC est perdu au cours de la dédifférenciation de
l’adénome ; puis une perte du gène p53 est observée lorsque le carcinome devient
invasif. L’ordre de survenue de ces mutations peut probablement varier selon les
tumeurs.
Au-delà, la cellule
tumorale acquiert des anomalies génétiques supplémentaires variables d’une
tumeur à l’autre, à la faveur de l’instabilité génétique provoquée par la perte
de p53.
Chez les individus
porteurs de mutations somatiques du gène APC et présentant une polypose
adénomateuse familiale, toutes les cellules somatiques de l’individu et
notamment les cellules épithéliales du colon ont franchi une 1re étape dans le
processus de carcinogenèse, qui se produit donc avec une fréquence beaucoup plus
élevée que dans la population générale et à un âge plus précoce.
4. Défaut de réparation
de l’ADN :
D’une manière générale,
tout phénomène biologique qui augmente le taux de mutation de l’ADN augmente le
risque de développer un cancer.
On a identifié ainsi
plusieurs syndromes de prédisposition au cancer qui résultent d’altération des
systèmes de réparation de l’ADN.
Le xeroderma pigmentosum
est un syndrome dans lequel les individus, le plus souvent des enfants,
présentent une anomalie d’un gène codant pour une des protéines impliquées dans
la réparation des lésions de l’ADN dues aux ultraviolets et vont présenter des
cancers cutanés multiples dans les 2 premières décennies de la vie.
Les cancers héréditaires
du côlon non polyposiques sont également liés à des altérations des gènes
(msh-2, mlh-1) d’un système de réparation de l’ADN (réparation des
mésappariements de l’ADN).
L’inactivation de ce
système entraîne l’apparition d’une instabilité génétique qui favorise
l’accumulation progressive de mutations.
L’instabilité génétique
observée dans ce syndrome prédispose aux cancers du côlon et à d’autres cancers
(endomètre, voies biliaires, estomac) à un âge précoce. D’autres mutations de
gènes impliqués dans la réparation et la réplication de l’ADN sont responsables
de maladies génétiques augmentant le risque de cancers comme le gène ATM dans
l’ataxie télangiectasie.
Chez les eucaryotes
supérieurs, dont l’homme, la majorité des cellules sont en G0 où elles exercent
leurs fonctions spécialisées.
L’entrée d’une cellule
dans le cycle cellulaire requiert la présence de facteurs de croissance dans le
milieu extracellulaire et de l’ancrage de la cellule à la matrice
extracellulaire par l’intermédiaire de molécules d’adhésion.
Les cellules cancéreuses
produisent fréquemment ces facteurs de croissance en excès, ou présentent une
activation permanente de leurs récepteurs et des voies de transduction du signal
en aval.
Les facteurs de croissance
sont des petites protéines de signalisation extracellulaire (cytokines) actives
à très faibles concentrations (10-9 à 10-10 mmol) qui vont induire l’entrée de
la cellule en phase S et parfois moduler la différenciation, la fonction et (ou)
la migration des cellules cibles exprimant leurs récepteurs spécifiques.
Ils agissent selon un
mécanisme paracrine, parfois autocrine et, plus rarement, endocrine.
Plus d’une centaine de
facteurs de croissance ont désormais été identifiés.
Leur nom provient des
circonstances de leur découverte ou de leur fonction EGF et PDGF.
Certains agissent sur une
très grande variété de cellules (EGF, PDGF), d’autres sur une population
cellulaire très limitée (interleukine 2, érythropoïétine).
Certains facteurs de
croissance, comme le TGF P (transforming growth factor (3) exercent des effets
biologiques opposés (effet prolifératif ou antiprolifératif) dans des types
cellulaires différents.
Plusieurs de ces facteurs
de croissance sont des proto¬oncogènes qui vont contribuer à l’acquisition d’un
phénotype cancéreux lorsqu’ils sont produits en excès (PDGF, EGF, macrophage
colony stimulating factor ou M-CSF) en agissant souvent selon un mécanisme
autocrine.
Ces facteurs de croissance
interagissent avec la portion extracellulaire des récepteurs transmembranaires
spécifiques présents à la surface de la cellule, parfois constitués d’une ou
plusieurs chaînes différentes.
Dans leur portion
intracellulaire, ces récepteurs, après interaction avec leur ligand, exercent
une activité enzymatique variable, tyrosine kinase (récepteur de l’EGF ou du
PDGF), sérine thréonine kinase (récepteur du TGF ), ou interagissent avec des
protéines G ancrées dans le feuillet interne de la membrane plasmique.
D’autres récepteurs sont
dépourvus d’activité enzymatique intrinsèque, mais s’associent à des protéines
cytosoliques qui vont transmettre directement le signal ; c’est le cas de
récepteurs d’interleukines (IL2, IL4, IL6), qui interagissent avec des kinases
(janus kinases ou Jak) qui vont phosphoryler et activer directement des facteurs
de transcription.
Les récepteurs tyrosine
kinases se « dimérisent » après interaction avec leur ligand, permettant ainsi
une autophosphorylation de tyrosine dans leur portion intracytoplasmique; les
récepteurs ainsi phosphorylés interagissent alors avec d’autres protéines
cytosoliques, qui activent la protéine ras, qui active elle-même d’autres
kinases intracytoplasmiques (c-raf). Par une cascade de phosphorylations, le
signal induit par le facteur de croissance est transmis jusqu’au noyau induisant
ainsi la transcription de certains gènes et permettant l’entrée dans le cycle
cellulaire.
Les facteurs de croissance
induisent l’expression de 2 grandes classes de gènes, des gènes à réponse
précoce (dont les proto-oncogènes c-fos et c-jun) et des gènes à réponse
retardée, qui sont activement transcrits au-delà d’une heure après l’exposition
de la cellule au facteur de croissance.
Parmi ces derniers
figurent les protéines de la machinerie de contrôle du cycle cellulaire,
c’est-à-dire les cyclines, et les protéines kinases dépendant des cyclines qui
permettent aux cellules de passer le point de restriction de fin de G1 et
d’entrer en phase S.
La protéine Rb joue un
rôle important dans ce phénomène : dans la cellule en G0, Rb se lie sous sa
forme déphosphorylée à des protéines régulatrices de la transcription qu’elle
inactive.
Après exposition à un
facteur de croissance, Rb va être phosphorylée, libérant ainsi ces facteurs
régulateurs de la transcription, permettant ainsi l’induction de la
transcription de gènes tardifs. Ainsi, 2 phénomènes biologiques aussi différents
que l’inactivation fonctionnelle de Rb par mutation des 2 copies du gène et
l’activation permanente d’une voie de signalisation d’un facteur de croissance
ont des conséquences assez similaires sur l’entrée d’une cellule dans le cycle
cellulaire.
Les facteurs de
croissance, les récepteurs de facteurs de croissance ou les protéines
cellulaires qu’ils activent en cascade sont des proto-oncogènes fréquemment
mutés (c-erbB-1, c-ras, c-raf), surexprimés (c-erbB-2), amplifiés (c-erbB-2)
dans les cellules cancéreuses induisant ainsi une activation permanente des
voies de transduction du signal indépendamment de la présence du ligand.
De ce fait, les cellules
cancéreuses ont une moindre dépendance aux facteurs de croissance que les
cellules normales.
La perte de l’inhibition
de contact résulte en fait d’une compétition entre les cellules poussant à haute
densité pour l’utilisation des facteurs de croissance présents en faible
quantité en culture.
Les cellules cancéreuses
affranchies de cette dépendance peuvent continuer à se diviser même en présence
de très faibles concentrations de ces facteurs de croissance dans le milieu de
culture.
Apoptose :
Le terme d’apoptose
désigne un processus actif de mort cellulaire programmée, au cours duquel la
cellule procède, par l’activation d’enzymes intracellulaires notamment, à une
destruction de son ADN et de certaines protéines conduisant à sa propre mort.
L’apoptose est un
processus essentiel pour l’homéostasie de l’individu, intervenant dans des
processus aussi divers que l’embryogenèse, la destruction des lymphocytes T
autoréactifs, la destruction des cellules en sénescence, des cellules infectées
par des virus, des cellules porteuses de mutations de l’ADN.
Sur le plan morphologique,
l’apoptose se traduit par une diminution de la taille de la cellule et de son
noyau, qui se fragmente.
L’ADN cellulaire est clivé
en fragments multiples de 200 paires de bases correspondant à la longueur du
brin d’ADN nucléosomique. Ce processus de dégradation des composants
intracellulaires est exécuté en partie par une catégorie d’enzymes particulières
appelées caspases dont on connaît désormais plus d’une dizaine de membres.
L’entrée en apoptose et
l’activation de ces caspases est également sous le contrôle d’une famille de
protéines intracellulaires situées dans les membranes d’organites
intracellulaires dont les mitochondries, les protéines de la famille bcl-2 ;
certaines de ces protéines induisent l’apoptose, d’autres l’inhibent.
Plusieurs types de signaux
entraînent la cellule à s’engager dans un processus d’apoptose. Des protéines
solubles ou transmembranaires (tumour necrosis factor alpha, TNF; ligand de Fas,
TRAIL) interagissent avec des récepteurs spécifiques à la surface de la cellule
cible (« récepteurs de mort ») qui activent la cascade des caspases dans la
cellule.
Leur mode d’action est
ainsi très voisin de celui des facteurs de croissance et ils peuvent d’ailleurs
parfois (TNF ) agir comme des facteurs de croissance et non pas d’apoptose pour
certains types cellulaires.
La privation de facteurs
de croissance entraîne l’apoptose de certains types cellulaires notamment de
cellules du système hématopoïétique.
Les lésions irréparables
de l’ADN, causées par des agents mutagènes comme les radiations ionisantes ou
les agents cytotoxiques utilisés en
chimiothérapie anticancéreuse entraînent
l’apoptose.
La protéine p53 normale
intervient dans ce phénomène. Les cellules cancéreuses présentent fréquemment
une altération des voies d’induction ou d’inhibition de l’apoptose.
La tumeur pour laquelle
une altération des voies de l’apoptose a été la mieux documentée est le lymphome
folliculaire, une tumeur maligne du lymphocyte B, caractérisé par la
translocation chromosomique spécifique t(14,18) : cette translocation juxtapose
le gène antiapoptotique bcl-2 avec les séquences régulatrices d’un gène des
immunoglobulines, entraînant ainsi une surexpression de bcl-2 dans le clone
cellulaire tumoral.
Cette translocation
confère au clone tumoral une survie prolongée.
Cela a pu être démontré
dans des lignées cellulaires où la surexpression de l’oncogène c-myc permet à la
cellule d’entrer en cycle mais la conduit en apoptose. La surexpression de bcl-2
avec c-myc dans la même lignée permet à la cellule d’échapper à l’apoptose et
lui confère un phénotype cancéreux.
Une surexpression de bcl-2
ou d’autres protéines antiapoptotiques de la même famille a désormais été
observée dans d’autres types de cancers notamment des carcinomes.
Cette surexpression est
parfois associée à une agressivité clinique particulière de la tumeur.
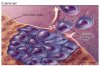
Dissémination tumorale :
L’une des caractéristiques de la
cellule tumorale est sa capacité
à envahir des régions de
l’organisme où elle ne devrait
pas se retrouver.
La capacité à disséminer dans
des tissus adjacents est la
première étape conduisant à
l’apparition de métastases.
La cellule cancéreuse, par
exemple d’origine épithéliale, a
ainsi la capacité de
s’affranchir de ses interactions
avec les cellules normales
environnantes, détruire la
membrane basale sous-jacente,
progresser à travers le tissu
conjonctif, franchir la paroi du
vaisseau lymphatique ou sanguin,
migrer dans la circulation
lymphatique ou sanguine,
franchir la paroi du capillaire
dans une autre région de
l’organisme, pour s’établir,
survivre et croître dans ce
nouvel environnement. Cette
succession d’étapes limitantes
nécessite l’acquisition par la
cellule cancéreuse de nouvelles
propriétés.
La perte de l’adhésion des
cellules tumorales aux cellules
normales adjacentes peut
résulter de plusieurs mécanismes
par exemple la perte de
l’expression de molécules
d’adhésion intercellulaire,
comme la cadhérine E, ou
l’inactivation fonctionnelle de
ces molécules d’adhésion après
exposition à des cytokines
telles que certains membres de
la famille des FGF. Dans les
lignées cellulaires dans
lesquelles l’expression de la
cadhérine E est perdue, la
transfection par la cadhérine E
permet de renverser le phénotype
métastatique, cette molécule
d’adhésion se comportant donc
comme un gène suppresseur de
tumeur.
L’étape suivante dans la
dissémination métastatique est
la dégradation des composants de
la matrice extracellulaire pour
permettre la progression de la
cellule tumorale dans le tissu
conjonctif environnant,
c’est-à-dire la membrane basale,
le chorion sous-jacent et la
paroi des vaisseaux sanguins ou
lymphatiques. Plusieurs familles
d’enzymes impliquées
physiologiquement dans le
remodelage du tissu conjonctif
jouent un rôle essentiel dans la
dégradation des protéines
fibreuses, des protéoglycanes et
des glycoprotéines de la matrice
extracellulaire : les héparanes,
les sérine, asparatyl et
cystéine-protéases, et les
métalloprotéases (matrix
metalloproteinase, MMP).
La cellule tumorale utilise les
MMP pour remodeler le tissu
extracellulaire environnant,
mais à la différence des
cellules normales, ce processus
s’associe à une motilité accrue
de la cellule tumorale, lui
permettant de progresser dans
les tissus et de les coloniser.
Pour progresser dans le tissu
conjonctif, les cellules
tumorales produisent divers
facteurs de motilité, des
cytokines (FGF1 et FGF2, IL6,
TGF …), des composants solubles
de la matrice extracellulaire
(fibronectine, laminine,
thrombospondine…) qui vont jouer
un rôle parfois opposé selon
qu’ils sont sous forme soluble
circulante ou insérés dans la
matrice conjonctive.
Seule la minorité des cellules
tumorales qui franchit la paroi
vasculaire et qui migre par la
circulation sanguine se fixe
dans un tissu à distance et
forme une métastase.
Dans la circulation, les
cellules tumorales interagissent
avec des plaquettes par
l’intermédiaire de molécules
d’adhésion (de la famille des
intégrines notamment). Pour
sortir du flux sanguin, les amas
de cellules tumorales et de
plaquettes interagissent avec
des molécules d’adhésion situées
au niveau des capillaires de
l’organe cible selon des
mécanismes probablement voisins
de ceux mis en jeu pour
l’extravasation des leucocytes.
L’amas de cellules tumorales et
de plaquettes induit la
production de médiateurs locaux
permettant une rétraction des
cellules endothéliales et
l’exposition de la membrane
basale.
La cellule tumorale pénètre la
matrice conjonctive de l’organe
cible en utilisant les facteurs
de motilité et les MMP, induit
une néo vascularisation avant de
proliférer pour donner une
métastase.
La relative spécificité des
organes sièges de métastases
pour un type de cancer donné est
encore mal comprise.
Il est certain que cette
spécificité résulte en partie de
phénomènes mécaniques: les
organes qui sont des points de
passage obligés du flux sanguin
pour certains organes (poumon,
foie) sont plus fréquemment le
siège de métastase.
Cependant, d’autres paramètres
interviennent probablement : des
molécules d’adhésion avec des
cellules tumorales spécifiques
des cellules endothéliales d’un
organe donné ont été décrites
dans certains modèles.
En outre, il est probable que
l’environnement tissulaire
normal de l’organe touché
(composants de la matrice,
facteurs de croissance produits
in situ) favorise le
développement de tel ou tel type
cellulaire dans un site
métastatique donné.
Enfin, l’un des paramètres
essentiels influençant la
croissance tumorale est la
capacité d’une cellule tumorale
à susciter le développement
d’une néo vascularisation
tumorale et, en particulier, la
différenciation et la croissance
de cellules endothéliales.
Ce processus, dénommé
angiogenèse, est indispensable à
la croissance tumorale au delà
d’un volume de quelques
millimètres cubes de tumeur dans
de nombreux modèles
expérimentaux.
La croissance tumorale est en
effet limitée au-delà de ce
volume par la disponibilité en
nutriments et en oxygène des
cellules tumorales.
La capacité d’une tumeur à
susciter une néovascularisation
semble indispensable au
développement de la tumeur, à la
fois localement et au site des
métastases.
On a ainsi pu décrire dans
certaines tumeurs (carcinomes in
situ du col de l’utérus) in
vivo, un état « prévasculaire »,
au cours duquel un petit amas de
cellules tumorales est présent
mais ne peut se développer au
delà d’un certain volume, et une
phase « vasculaire » où la
tumeur développe une
néovascularisation et peut
croître et éventuellement
disséminer.
Des mécanismes similaires sont
mis en jeu pour le démarrage de
la croissance de métastases «
dormantes » dans des modèles
animaux et peut être chez
l’homme.
Le passage d’une phase
prévasculaire à une phase
vasculaire est lié à la
production, le plus souvent par
les cellules tumorales
elles-mêmes, de facteurs de
croissance des cellules
endothéliales, appelés facteurs
angiogéniques.
Ces facteurs de croissance sont
des cytokines qui sont parfois
également des facteurs de
croissance autocrines pour les
cellules tumorales elles-mêmes.
Parmi ces facteurs on peut
citer, le vascular endothelial
growth factor ou VEGF,
l’angiogénine, les FGF1, 2 et 4,
le TGF , l’hepatocyte growth
factor ou HGF.
Ces cytokines produites par les
cellules tumorales exercent
ainsi fréquemment des propriétés
biologiques multiples et
interviennent à des étapes
différentes de la progression
tumorale, modulant aussi bien la
croissance de la cellule, que sa
motilité et sa capacité à
induire une néovascularisation.
La compréhension des mécanismes
biologiques de la dissémination
tumorale permet désormais
d’envisager des stratégies
autres que les traitements
cytotoxiques pour le traitement
des tumeurs disséminées.
Actuellement, des facteurs
inhibiteurs des métalloprotéase,
et de l’angiogenèse sont en
expérimentation clinique.
Incidence pronostique :
La meilleure compréhension des
mécanismes biologiques de la
cancérogenèse et notamment des
altérations génétiques des
cellules tumorales a conduit à
de nouvelles classifications des
tumeurs humaines.
Certaines maladies néoplasiques
sont désormais identifiées sur
la base d’une altération
génétique spécifique
(translocation) autant que sur
l’analyse cytologique ou
histologique classique : c’est
le cas de la leucémie myéloïde
chronique et de la t(9,22), des
sarcomes d’Ewing et de la t(11,
22) et ses variantes, des
lymphomes du manteau et de la
t(11,14).
Cette nosologie moléculaire des
maladies a parfois permis
d’individualiser des groupes de
patients avec un pronostic
complètement différent d’autres
patients porteurs de maladies
voisines ou similaires sur des
bases cytologiques histologiques
ou phénotypiques : les lymphomes
du manteau caractérisés par leur
t(11,14) ont un pronostic moins
bon que les patients porteurs
d’autres lymphomes diffus B à
petites cellules, les leucémies
aiguës non lymphoblastiques de
l’enfant avec t(9,22) ou une
translocation en 11q23 sont
associées à un pronostic
défavorable. En outre au sein
d’un même groupe de maladies
néoplasiques, par exemple les
adénocarcinomes du sein,
l’altération de l’expression
d’un gène spécifique est
corrélée à un pronostic
particulier, souvent moins bon.
C’est le cas pour les mutations
inactivatrices de gènes
suppresseurs de tumeur (p53 ou
Rb pour des cancers du sein,
lymphomes, leucémies) pour la
surexpression de proto-oncogènes
codant pour des tyrosine kinases
récepteurs de facteurs de
croissance (c-erbB2, dans les
cancers du sein dans certains
sarcomes), pour l’amplification
de proto-oncogènes codant pour
des facteurs de transcription
(N-myc dans le neuroblastome),
pour la production in vivo de
facteurs de croissance (VEGF,
IL6).
Ces paramètres biologiques ne
sont pas pour la plupart étudiés
en routine à l’heure actuelle.
La présence de ces anomalies
moléculaires semble cependant
influencer la réponse à certains
traitement antinéoplasiques
(efficacité des cytotoxiques) et
il est donc possible qu’ils
soient plus largement utilisés
dans les années à venir.
En outre, le développement de
traitements spécifiques dirigés
contre ces oncogènes (par
exemple, un anticorps monoclonal
anti-erb-B2) nécessitera
probablement une analyse
systématique de sa sur
expression.
Enfin, la mise à disposition
d’outils tels que les puces à
ADN, permettant l’analyse de
milliers de gènes dans une
tumeur donnée et ainsi une
analyse étendue des anomalies
génétiques de milliers de
tumeurs, va probablement
bouleverser dans les années à
venir à la fois la
classification nosologique de
ces maladies et conduire à
identifier des sous-groupes
pronostiques et thérapeutiques
encore insoupçonnés
actuellement.
Conclusion :
• Les proto-oncogènes sont des
gènes normaux qui vont subir des
mutations activatrices dans les
cellules cancéreuses, les
transformant ainsi en oncogènes
capables d’induire une division
cellulaire incontrôlée.
• Ils codent souvent pour des
protéines impliquées dans la
transmission du signal
mitogénique de la membrane
plasmique au noyau. Leur
mutation induit généralement
leur activation permanente,
rendant les cellules moins
dépendantes aux facteurs de
croissance exogènes pour
l’entrée en mitose.
• Les gènes suppresseurs de
tumeurs ont pour rôle
physiologique de freiner la
division cellulaire, lorsque la
cellule n’est pas dans une
situation adéquate (manque de
facteur de croissance, mutations
de l’ADN). Leur destruction
permet à la cellule cancéreuse
d’entrer en division,
d’accumuler des anomalies
génétiques supplémentaires ou
d’échapper aux signaux
entraînant l’apoptose.
• La surexpression de protéines
anti-apoptotiques est l’un des
mécanismes moléculaires
conduisant à la cancérogenèse
dans certains modèles.
• La transformation d’une
cellule normale en cellule
cancéreuse fait intervenir
plusieurs des mutations
consécutives de gènes
suppresseurs de tumeurs et
d’oncogènes. Chez la plupart des
individus, ces altérations
génétiques de la cellule
tumorale sont acquises.
Cependant, dans certains
syndromes de cancers familiaux,
les individus atteints
présentent une mutation
somatique qui inactive un gène,
généralement un gène suppresseur
de tumeur. Les individus
affectés présentent ainsi une
incidence accrue de certaines
tumeurs qui surviennent en outre
à un âge précoce.
• Les facteurs de croissance
sont fréquemment produits en
excès par les cellules
cancéreuses. Ils interviennent à
plusieurs étapes dans la
progression du cancer, agissant
comme facteur anti-apoptotique,
facteur de croissance, facteur
de motilité cellulaire ou comme
facteurs angiogéniques.
• La dissémination des cellules
tumorales, à travers la membrane
basale, dans le tissu
conjonctif, à travers la paroi
des vaisseaux et dans les sites
métastatiques fait intervenir
l’altération de la structure ou
de la fonction des molécules
d’adhésion à leur surface, la
production d’enzymes capables de
détruire les fibres et les
glycoprotéines de la matrice, la
production en excès de cytokines
agissant comme des facteurs de
motilité, la production de
néovaisseaux capables de
vasculariser la tumeur primaire
ou la métastase en croissance.
• La caractérisation des
anomalies moléculaires des
cellules cancéreuses permet la
mise en place d’une nouvelle
classification de ces maladies
reposant sur des critères
moléculaires. Certaines
anomalies génétiques constituent
en outre un facteur pronostique
pour certaines affections
néoplasiques.