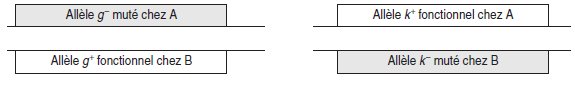La définition fonctionnelle du gène : la découverte de la relation un gène/une enzyme
La fonction du gène, c’est-à-dire son contenu informatif est demeuré mystérieux
jusque dans les années 1940, quand des expérimentations, notamment sur des
mutants simples (touchés en un seul gène) du métabolisme chez la levure, ont permis
de montrer qu’un gène codait pour une chaîne peptidique (le dogme originel « un
gène-une enzyme » ayant été rapidement remplacé par la formule plus générale « un
gène-une chaîne peptidique »).
La levure, comme tous les microorganismes, est capable d’assurer la biosynthèse
de toutes les molécules organiques simples de la biochimie, notamment les acides
aminés.
Un mutant incapable de produire une telle molécule est dit auxotrophe, alors
que la souche sauvage, capable de la biosynthétiser est dite prototrophe. Un mutant
auxotrophe peut néanmoins pousser s’il est mis en culture sur un milieu minimal
(une source de carbone et d’azote, et une série d’éléments minéraux) additionné de
la molécule qu’il est incapable de produire, mais qu’il va trouver dans ce milieu.
Disposant d’une série de mutants auxotrophes pour un acide aminé (comme
l’histidine), les généticiens ont pu montrer que de tels mutants pouvaient aussi
pousser sur des milieux minimum additionnés d’un des intermédiaires ou précurseurs
de la chaîne de biosynthèse de l’acide aminé.
Le concept de chaîne métabolique gouvernée par l’action des gènes était attesté
par plusieurs faits :
– certains mutants, poussant sur un milieu additionné de l’acide aminé terminal,
excrétaient une molécule précurseur de celui-ci, ce qui montrait bien que la chaîne
était bloquée à l’étape de transformation de ce précurseur-intermédiaire;
– les différents mutants ne pouvaient pas pousser avec n’importe lequel des précurseurs,
ce qui montrait bien qu’ils n’étaient pas bloqués dans la même étape de la
chaîne de biosynthèse;
– certains mutants incapables de pousser avec un précurseur A se révélaient capables
de pousser avec un précurseur B, ce qui prouvait que l’ordre de biosynthèse
était bien A-B-acide aminé terminal.
Grâce à ce type d’expérimentations les généticiens purent non seulement établir
ou préciser les différentes chaînes métaboliques, mais surtout arriver à la conclusion
qu’un mutant auxotrophe, muté dans un gène, était, en général, incapable d’assurer
une et une seule des étapes d’une chaîne de biosynthèse.
De là, ils conclurent que
chaque chaîne de biosynthèse était dépendante d’un ensemble de gènes, chacun de
ces gènes gouvernant la réalisation d’une étape.
Puis on a montré comment un gène gouvernait une étape d’une chaîne de biosynthèse
en montrant que son contenu informatif spécifiait précisément l’enzyme
permettant la réalisation de cette étape.
La démonstration est venue du fait que
certains mutants étaient dépourvus d’une enzyme connue pour intervenir dans la
chaîne étudiée. D’autres arguments s’ajoutèrent, notamment grâce aux études des
pathologies héréditaires de l’hémoglobine chez l’homme.
Dans la plupart des cas,
ces pathologies héréditaires résultent de la mutation d’un seul gène (car se transmettant
selon un mode mendélien, ségrégation 2 × 2), et sont caractérisées par la
présence d’une hémoglobine anormale dont la chaîne α (ou β, selon les cas) diffère
systématiquement de la chaîne normale par un seul et même acide aminé.
Cette observation attestait non seulement l’idée que le gène était bien un message
codant une chaîne peptidique (α ou β), mais aussi l’idée que ce type de mutation
touchait ponctuellement le message génétique puisque tous les acides aminés, sauf
un, étaient correctement spécifiés et ordonnés, ce qui conduisait à l’hypothèse,
confirmée par la biologie moléculaire, que le gène spécifie, par sa séquence, la
nature et l’ordre des acides aminés du produit peptidique constituant son « produit ».
Depuis cette date le concept de gène est devenu beaucoup plus complexe .
La
complémentation fonctionnelle et le test d'allélisme :
La complémentation fonctionnelle est un phénomène qui a permis de comprendre la
fonction biochimique du gène car il trouve son explication dans cette fonction.
C’est
un phénomène résultant du fait qu’un gène code pour une chaîne peptidique, un
produit diffusible au sein de la cellule (voire au sein de l’organisme si elle est
sécrétée).
D’un point de vue expérimental, la complémentation fonctionnelle permet de
comprendre pourquoi et comment le croisement de deux souches mutantes de même
phénotype peuvent avoir une descendance de phénotype sauvage, en considérant
l’exemple suivant, où il est très important de noter l’enchaînement des étapes expérimentales
et leur relation logique.
On dispose de trois souches haploïdes de levure, respectivement notées A, B et C,
auxotrophes pour l’histidine, phénotype noté [his–], et de la souche SSR prototrophe
pour l’histidine, de phénotype noté [his+].
On souhaite, par l’analyse génétique de
ces mutants, entreprendre l’analyse génétique du processus de biosynthèse de
l’histidine.
A - Croisement des mutants par la souche sauvage SSR :
test de dominance/récessivité
:
Le croisement de chacune des souches A, B ou C par la souche SSR, donne des
cellules diploïdes de phénotype [his+]. Ce test de dominance montre que les trois
phénotypes d’auxotrophie sont récessifs.
L’interprétation fonctionnelle du caractère
récessif de l’auxotrophie est facile si on considère la fonction d’un gène :
– la souche SSR possède, pour un gène g, un allèle fonctionnel, noté g+, qui lui
permet de produire une des enzymes de la chaîne de biosynthèse de l’histidine;
– la souche A est mutée dans ce gène g, elle possède un allèle muté, noté g–, non
fonctionnel, soit parce que l’enzyme est absente, soit parce qu’elle est présente,
mais non fonctionnelle (tout dépend de l’effet biochimique de la mutation).
La
chaîne de biosynthèse de l’histidine est alors bloquée au niveau de l’étape
gouvernée par l’enzyme résultant de l’expression du gène g;
– la souche diploïde, issue du croisement SSR × A, possède deux exemplaires du
gène g, un exemplaire sauvage g+ venant du parent SSR et un allèle muté g–
venant du parent A. Cet hétérozygote g+//g– peut se schématiser ainsi :
– dans la cellule diploïde, l’expression de l’allèle g+ du gène g conduit à la présence
d’une enzyme fonctionnelle permettant de réaliser l’étape bloquée chez le mutant A.
Si la quantité produite d’enzyme est suffisante pour que cette étape puisse être
réalisée sans problème, la présence de l’allèle muté g– n’a aucune conséquence
phénotypique et le phénotype du diploïde est sauvage [his+].
Il est capable
d’assurer la biosynthèse de l’histidine.
L’effet de l’allèle sauvage est dominant s’il compense celui de l’allèle muté (pour
plus de détails, voir 5.5).
La même interprétation vaut pour les souches mutantes B ou C, mais rien ne
permet de dire que A, B ou C sont mutées dans un même gène ou dans des gènes
différents !
Par ailleurs, rien ne permet de dire que A, B ou C ne sont mutées que dans un seul
des gènes de la chaîne de biosynthèse, information qui résultera de l’étape suivante
de l’analyse génétique.
B - Analyse génétique de la méiose chez les diploïdes
issus du croisement mutant × SSR :
Il s’agit ici de tester la ségrégation 2 × 2 des phénotypes [his+] et [his–], chez les spores
haploïdes résultant de la méiose des diploïdes issus des croisement SSR × mutant.
S’il y a ségrégation 2 × 2, on peut conclure que la souche mutante ne
diffère de la souche sauvage que pour un seul gène, la méiose donnant les proportions
attendues dans le cas d’un seul couple d’allèles.
Si la ségrégation n’est pas une ségrégation 2 × 2, on doit conclure que la souche
mutante diffère de la souche sauvage pour plus d’un gène (en l’occurrence, elle est
bloquée dans plusieurs étapes) et il convient, quand c’est possible, de dénombrer le
nombre de gènes mutés, par l’analyse des fréquences des différents phénotypes.
Considérons, dans notre exemple que la ségrégation 2 × 2 étant observée chez les
trois types de diploïdes, on puisse conclure que chaque souche A, B ou C n’est
mutée que dans un seul des gènes de la chaîne de biosynthèse de l’histidine.
C - Croisements entre souches mutantes :
test de complémentation fonctionnelle et test d’allélisme :
Le test de complémentation fonctionnelle est le moyen expérimental qui permet de
montrer que deux souches mutantes sont ou ne sont pas mutées dans un même gène,
ou dans le même gène, si on a montré par ailleurs que chacune des souches n’était
mutée que dans un seul gène (ségrégation 2 × 2 dans un croisement avec la SSR, voir
plus haut).
Si on procède au croisement de deux souches mutantes de phénotypes [his–], on
peut s’attendre, compte tenu de ce qu’on sait de la fonction d’un gène et de notre
interprétation de la récessivité des phénotypes mutants, à deux résultats possibles :
– soit les deux souches sont mutées dans des gènes différents, alors le phénotype du
diploïde issu du croisement entre elles est sauvage [his+];
– soit les deux souches sont mutées dans le même gène, alors le phénotype du
diploïde issu du croisement entre elles est muté [his–].
Ces deux types de résultats sont faciles à comprendre à partir des figures
suivantes.
Cas 1. Les souches A et B ne sont pas mutées dans le même gène, A est mutée
dans un gène g et B est mutée dans un gène k (fig. 5.1).
Figure 5.1 Représentation schématique d’un diploïde
formé d’apports haploïdes mutés dans deux gènes différents.
Dans la cellule diploïde, l’expression de l’allèle g+ du gène g conduit à la présence
d’une enzyme fonctionnelle permettant de réaliser l’étape bloquée chez le mutant A,
comme cela se passait chez le diploïde issu du croisement mutant A × SSR.
De
même l’expression de l’allèle k+ du gène k conduit à la présence d’une enzyme fonctionnelle
permettant de réaliser l’étape bloquée chez le mutant B, comme cela se
passait chez le diploïde issu du croisement mutant B × SSR. Il y a double compensation
allélique.
Au total, les deux étapes bloquées, l’une chez A, l’autre chez B, sont désormais
réalisables : il y a complémentation fonctionnelle.
Chacune des souches, étant mutée
dans un gène différent, apporte, chez le diploïde issu de leur croisement, la fonction
dont l’autre est dépourvue, et le diploïde est alors de phénotype [his+], contrairement
aux deux souches parentales.
D’un point de vue expérimental, l’observation de la complémentation fonctionnelle
permet de conclure que les deux souches mutantes étudiées, ici A et B, ne sont
pas mutées dans le même gène. Le test de complémentation fonctionnelle est donc
aussi un test d’allélisme puisqu’on peut conclure que les mutations g– et k– des
souches A et B ne sont pas alléliques car elles ne touchent pas le même gène.
Cas 2. Les souches A et C sont mutées dans le même gène g (fig. 5.2).
Figure 5.2 Représentation schématique d’un diploïde
formé d’apports haploïdes mutés dans le même gène.
Chez ce diploïde, les deux exemplaires du gène g sont mutés et non fonctionnels;
il ne peut, pas plus que chacune des souches parentales, réaliser l’étape bloquée par
l’absence de l’enzyme codée par le gène g (ou sa présence, mais dans un état non
fonctionnel).
Il n’y a pas complémentation fonctionnelle parce que les deux souches A
et C sont mutées dans le même gène, le diploïde est de phénotype [his–], comme les
souches parentales.
D’un point de vue expérimental, le test de complémentation fonctionnelle est
donc aussi un test d’allèlisme puisqu’on peut conclure que les mutations des souches A
et B sont alléliques puisqu’elles touchent le même gène.
En conséquence, on doit s’attendre à observer de la complémentation fonctionnelle
dans le croisement B × C, puisque A est muté dans un gène différent de B et
que A est muté dans le même gène que C.
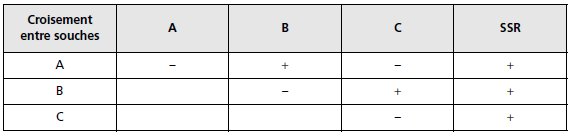
À l’issue d’un test de complémentation
fonctionnelle, les résultats peuvent se présenter sous la forme d’un tableau (tabl. 5.1),
où la dernière colonne rappelle les résultats du test de dominance, où les signes « – »
et « + » indiquent, respectivement, l’incapacité de produire de l’histidine (pas de
complémentation fonctionnelle), ou la capacité d’en produire (complémentation
fonctionnelle ou compensation allélique).
TABLEAU 5.1 TABLEAU DE RÉSULTATS DE TESTS DE COMPLÉMENTATION ET DE DOMINANCE.
Remarque 1. L’interprétation précédente n’est valable que parce qu’on sait
que chacune des souches A, B ou C n’est mutée que dans un seul gène.
Imaginons
que C soit un double mutant (pas de ségrégation 2 × 2 chez le diploïde
issu de la méiose C × SSR), on pourrait observer d’autres résultats (tabl. 5.2).
TABLEAU 5.2 RÉSULTAT D’UN TEST DE COMPLÉMENTATION FAISANT APPARAÎTRE UN DOUBLE MUTANT.
L’interprétation conduirait à considérer que A et C sont mutés dans un même
gène (on ne peut pas dire le même gène puisque A est un mutant simple et que
C est un mutant double) et que B et C sont aussi mutés dans un même gène,
mais différent du précédent, puisque A et B complémentent.
Remarque 2. L’interprétation d’un test de complémentation fonctionnelle
n’est possible que pour les croisements entre deux mutants récessifs.
Si parmi des mutants de même phénotype, certains sont dominants (pas de
compensation allélique de l’allèle sauvage), ce que permet de déterminer le
test de dominance par le croisement avec la SSR, ces mutants dominants ne
peuvent donner lieu à aucune analyse fonctionnelle dans des croisements avec
d’autres mutants (dominants ou même récessifs).
Il suffit de reprendre les
figures 5.1 et 5.2 et les raisonnements associés pour voir que le phénotype du diploïde sera toujours muté, que les deux souches parentales soient ou ne
soient pas mutées dans le même gène.
C’est pourquoi on a souvent l’habitude, dans un tableau de croisements entre
mutants, d’y ajouter les croisements mutant × SSR, ce qui permet d’exclure
de l’analyse tout mutant dominant.
Remarque 3. Deux souches mutées dans un même gène, comme A et C dans
l’exemple ci-dessus, ne sont pas forcément mutées, ni au même site dans le
gène, ni de la même manière (il y a plusieurs types différents de mutation d’un
gène, mutation stop ou non-sens, mutation faux-sens, mutation de décalage du
cadre de lecture, mutation d’épissage, délétion ou insertion de triplets ou de
séquences plus longues, mutation dans le promoteur, etc.).
Deux allèles d’un gène sont dits homoallèles quand ils sont porteurs de la même
mutation (même site, même type de mutation), deux allèles d’un gène sont dits
hétéroallèles quand ils sont porteurs de deux mutations différentes de ce gène
(mutations du même gène mais en un site différent et/ou d’un type différent).
Mais, dès lors qu’un diploïde est porteur de deux exemplaires mutés non fonctionnels
d’un gène, il est, du point de vue fonctionnel, incapable de réaliser
l’opération gouvernée par ce gène, que les deux exemplaires soient homoalléliques
ou hétéroalléliques.
Un test de complémentation fonctionnelle, s’il permet de conclure que A et B
sont mutés dans le même gène, ne permet pas de conclure sur la nature homoallélique ou hétéroallélique des mutations touchant A et B.
On peut
cependant dire, si les mutants A et B ont été obtenus indépendamment, que la
plus grande probabilité est qu’il s’agisse d’hétéroallèles.
Un diploïde porteur de deux hétéroallèles a un génotype dit « hétérozygote
composite » et un phénotype muté, puisqu’il n’y a pas de complémentation
fonctionnelle entre mutations homo et/ou hétéroalléliques.