De Colomb ou Magellan à La Perouse, tous les grands capitaines ont embarqué à
leur bord cartographes et géomètres susceptibles de donner une forme, au moins un
contour à toutes les terra incognita que leurs découvertes ajoutaient peu à peu aux
cartes maritimes et à celles des empires.
C’est un principe général auquel n’échappent ni les gènes ni le génome que
l’étude d’un objet commence par la description de sa forme et de sa position dans
l’espace.
Aussi, bien avant de savoir quelle est la fonction d’un gène et comment les
produits de plusieurs gènes pouvaient interagir dans la réalisation d’un phénomène,
le généticien s’est préoccupé de localiser ces gènes afin d’obtenir une vision cartographique
du génome de l’espèce étudiée.
La cartographie des gènes est essentielle car elle facilite ultérieurement les
analyses génétiques, y compris les analyses fonctionnelles; aujourd’hui beaucoup de
recherches qui ont trait au cancer se font sur des gènes de levure ou de drosophile,
homologues à des gènes humains, tout simplement parce que ces organismes sont
simples à cultiver en grand nombre, à croiser avec des temps de générations courts,
mais aussi parce que leur génome est particulièrement bien cartographié.
Les problèmes cartographiques que le généticien rencontre sont de plusieurs ordres :
– la localisation ou assignation chromosomique;
– la liaison physique et génétique entre gènes d’un même chromosome, avec estimation
de leurs distances respectives;
– la carte fine d’un gène c’est-à-dire la répartition des différents sites des mutations
connues du gène et, éventuellement, leurs distances respectives, physiques en
nombre de nucléotides, ou génétiques en fréquence de recombinaison.
Il convient de remarquer que les distances physiques ne sont pas des distances
génétiques car la probabilité de crossing-over n’est pas constante le long de l’ADN,
contrairement à l’hypothèse simple de départ d’une analyse génétique.
En effet des
« points chauds » de recombinaison vont donner une grande distance génétique
entre deux sites physiquement proches alors que des séquences où les crossing-over
sont inhibés donneront des distances génétiques très petites entre des points physiquement
très éloignés.
Deux exemples le montrent facilement.
Les distances physiques sont les mêmes
sur des chromosomes humains portés par un organisme masculin ou féminin, mais il
y a deux fois moins de crossing-over chez l’homme que chez la femme et les
distances génétiques sont toujours divisées par deux dans le premier sexe; chez la
drosophile, la situation est extrême puisque l’absence de crossing-over chez le mâle
aboutit, dans ce sexe, à l’absence de gamètes recombinés par crossing-over, donc à
des distances génétiques nulles.
L’association
ou localisation chromosomique :
L’étude génétique de plusieurs gènes permet de définir des groupes de liaison, c’est-à-dire
des ensembles de gènes génétiquement liés entre eux sur un même chromosome,
mais seule la localisation d’au moins un gène de ce groupe sur un des chromosomes de
l’espèce peut permettre d’attacher ce groupe de liaison à un chromosome spécifique.
L’enjeu de la localisation chromosomique des gènes consiste précisément à identifier
lequel des chromosomes est porteur d’un gène donné, ce qui permettra d’y
localiser tous les gènes du même groupe de liaison.
La localisation des gènes du chromosome X, chez les organismes pourvus d’un
système hétérosomique, est facilement résolue puisque les phénotypes dépendant de
ces gènes présentent une transmission héréditaire typique.
De nombreuses méthodes spécifiques ont été développées pour l’assignation
chromosomique d’un gène, associant, selon les cas, l’analyse génétique, la cytogénétique,
c’est-à-dire la visualisation des chromosomes (caryotype) ou la biologie
moléculaire.
• L’association d’un phénotype résultant de la mutation d’un gène et d’une délétion
chromosomique permet de conclure que ce gène est sans doute localisé sur ce chromosome,
dans la partie délétée, ce qui entraîne une mutation par perte de fonction.
D’autres anomalies structurales du chromosome, comme une duplication ou une
inversion, peuvent aussi être utilisées pour assigner un gène, et même, plus finement,
localiser son emplacement sur le chromosome.
• L’utilisation de la séquence clonée d’un gène comme sonde pour réaliser une
hybridation in situ sur caryotype (FISH, fluorescent in situ hybridization) permet de
visualiser, grâce au signal de marquage de la sonde, la localisation chromosomique et même son emplacement plus ou moins précis sur le chromosome (bras court ou
bras long, position par rapport aux bandes d’intensité en coloration différentielle).
• L’utilisation de lignées cellulaires stables hybrides homme-rongeur est aussi un
outil d’assignation et de localisation chromosomique d’un gène humain.
Ces
lignées, cultivées in vitro, ont gardé la totalité des chromosomes rongeurs mais n’ont
gardé qu’un petit nombre de chromosomes humains.
Elles permettent de réaliser
l’assignation ou la localisation chromosomique d’un gène humain :
– soit par l’étude de son expression qui sera confinée aux seules lignées ayant
conservé le chromosome porteur du gène (cela suppose que le produit du gène
humain puisse être dosé);
– soit que le gène humain, ayant été cloné, sa séquence soit utilisée comme sonde
pour hybrider un Southern blot de fragments des différentes lignées, ce qui donnera
un signal positif dans les seules lignées ayant conservé le chromosome porteur
du gène.
On a donné le nom de « génétique somatique » à ce type d’analyses génétiques
fondées sur l’étude de lignées somatiques en culture, et non de croisements.
• Un autre type de stratégie consiste à faire une « revue génomique » (génome scan).
Cette stratégie, définie par Botstein chez l’homme, consiste à étudier systématiquement
la liaison génétique éventuelle entre le gène à assigner et un ensemble de
marqueurs génétiques (RFLP ou VNTR) répartis et cartographiés sur les différents
chromosomes.
En cas d’indépendance génétique entre le gène étudié et un marqueur, la zone
génomique autour du marqueur est exclue comme zone possible de localisation du
gène.
La revue génomique continue jusqu’à la découverte d’un marqueur lié au
gène, ce qui permettra automatiquement la localisation du gène dans le voisinage du
marqueur, sur le chromosome où est localisé ce marqueur.
On dispose pour assigner une séquence identifiée d’ADN, de nouveaux outils,
apparus depuis une dizaine d’années, les chromosomes artificiels de levure (YAC :
Yeast Artificial Chromosome) ou de bactérie (BAC : Bacterial Artificial Chromosome)
et du nombre croissant de génomes ayant été séquencés. Mais l’ensemble des stratégies
d’assignation avec ces outils déborde le cadre de cet ouvrage.
La
cartographie par analyse de liaison génétique :
Le but du jeu est ici d’étudier plusieurs gènes ou marqueurs génétiques afin de
statuer sur leur indépendance ou leur liaison génétique.
Dans ce dernier cas, une
distance peut être estimée et les valeurs respectives des distances entre gènes ou
marqueurs d’un même groupe de liaison peuvent permettre de les cartographier, de
les ordonner sur l’axe formé par le chromosome.
Le moyen expérimental le plus connu et le plus ancien est l’étude de la recombinaison
génétique lors de la méiose mais la génétique a développé de
nombreuses techniques dont certaines sont évoquées plus haut.
La
cartographie par délétion :
L’existence de délétions, ou leur induction favorisée par mutagenèse aux rayons X
ou d’autres radiations, peut être un outil efficace de cartographie des gènes ou, à une
échelle plus fine, des sites de mutation dans un gène (voir exercices d’application).
Dans la mesure où une délétion est une perte de matériel génétique, elle constitue
le plus souvent, pour les gènes touchés, une mutation de perte de fonction, en
général récessive, ce qui inclut souvent les mutants par délétion dans les analyses de
complémentation fonctionnelle.
Il convient de remarquer qu’une délétion n’est pas systématiquement une perte de
fonction; elle peut, par exemple, rendre un gène constitutif si elle touche son seul
site de fixation d’un répresseur, ou si elle fusionne sa séquence codante avec le
promoteur d’un autre gène.
A - Cartographie par délétion des sites de mutation d’un gène
:
Si on dispose de plusieurs mutants ponctuels du même gène et, par ailleurs, de
plusieurs mutants par délétion de ce gène, il est alors possible, par les croisements
entre mutants, d’ordonner les sites de mutations et symétriquement de délimiter
l’amplitude des délétions (voir exercices d’application).
Dans tous les cas, la stratégie expérimentale consiste à croiser entre eux deux
mutants A et B de même phénotype dont l’un, A, est porteur d’une mutation par délétion
partielle du gène (du moins on l’espère) alors que l’autre, B, est porteur d’une
mutation ponctuelle.
La question cartographique posée est alors la suivante, le site
de mutation ponctuelle dans le gène étudié, fourni par B, est-il dans ou hors de la
zone génomique délétée chez A ?
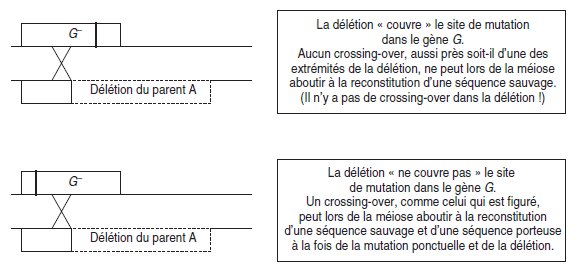
Figure 6.1 Conséquences génétiques de la recombinaison par crossing-over selon
que la délétion couvre ou ne couvre pas le site de mutation ponctuelle sur la séquence
homologue du gène (seules deux chromatides homologues non soeurs ont été figurées).
Dans le premier cas, on dit que le site de mutation ponctuelle est couvert par la
délétion, dans le second cas, qu’il ne l’est pas, ce qui a des conséquences opposées,
du point de vue de la recombinaison génétique lors de la méiose (fig. 6.1).
En croisant plusieurs mutants ponctuels avec le même mutant par délétion, on
peut classer les mutations ponctuelles en deux groupes, celles qui sont « couvertes »
et celles qui ne le sont pas, qui lui sont extérieures, ce qui est une façon de les
ordonner.
En opérant de la même manière avec des mutants porteurs d’autres délétions
partielles, on peut alors ordonner les sites de mutations les uns par rapport aux autres
et définir, en même temps, les limites de chaque délétion partielle dans le gène
étudié (voir exercices).
B - Différences entre mutants par délétion
et mutants ponctuels multiples :
Lorsque deux gènes sont contigus et que des mutations dans l’un ou l’autre conduisent
à un même phénotype, il est possible de le mettre en évidence si des mutants
simples (mutés dans un seul gène) et récessifs sont capables de donner par croisement
un diploïde sauvage (complémentation fonctionnelle).
Des doubles mutants
ponctuels, comme des mutants par délétion couvrant les deux gènes, apparaîtront
fonctionnellement semblables, puisqu’appartenant simultanément aux deux groupes
de complémentation.
Il est pourtant possible de les différencier dans la mesure où ils
peuvent présenter des propriétés génétiques différentes :
– la délétion étant un seul événement mutationnel, on observera une stricte ségrégation
2/2 chez le diploïde issu du croisement entre le mutant par délétion et le
sauvage.
En revanche, on pourra éventuellement exclure la ségrégation 2/2 chez le
diploïde issu du croisement entre sauvage et le double mutant, à condition
d’observer un nombre de méioses suffisamment élevé pour que les crossing-over,
entre les sites, donnent un surplus significatif de gamètes recombinés simples
mutés (fig. 6.2).
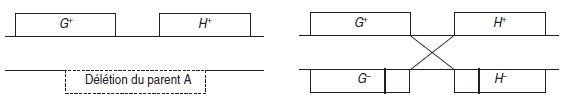
Figure 6.2 Conséquences génétiques de la méiose selon que deux gènes contigus
sont mutés par une délétion chevauchante ou deux mutations ponctuelles.
Le premier diploïde donnera strictement 1/2 de gamètes sauvages et 1/2 de gamètes
mutés. Le deuxième diploïde donnera (1 – r)/2 de gamètes sauvages et (1 + r)/2 de
gamètes mutés simples ou doubles; r étant le taux de recombinaison entre les sites
mesurable significativement si le nombre de gamètes étudiés, c’est-à-dire de méioses,
est élevé (seules deux chromatides homologues non soeurs ont été figurées).
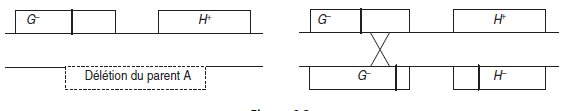
– le double mutant, croisé avec un mutant ponctuel simple peut donner, à la méiose
du diploïde, des gamètes sauvages, alors que le mutant par délétion n’en donnera
jamais, si la mutation ponctuelle est couverte (fig. 6.3).
Figure 6.3.
Le premier diploïde ne donnera jamais de gamètes sauvages. Le deuxième diploïde
donnera quelques gamètes sauvages si le nombre de gamètes étudiés, c’est à dire de
méiose, est élevé (seules deux chromatides homologues non soeurs ont été figurées).