L’analyse génétique d’un caractère chez un organisme suppose que celui-ci présente
une variabilité qui permette de définir au moins deux formes de ce caractère, désignées
comme phénotypes. En général cette analyse génétique (on dit même parfois
« dissection génétique » du caractère ou du phénomène biologique) consiste, après
avoir défini un phénotype de référence encore appelé « phénotype sauvage », à isoler
le plus grand nombre de « mutants » présentant une variation phénotypique héréditairement
stable.
On entreprend alors l’analyse génétique des mutants par des croisements avec la
souche sauvage de référence (SSR) afin d’établir la dominance ou la récessivité des
phénotypes mutants, prélude à la mise en oeuvre des tests de complémentation, par
les croisements entre mutants récessifs, qui permettront, avec les tests de ségrégation
des F1 issus des croisements entre mutants et SSR, de regrouper les différents
mutants et de dénombrer ainsi les gènes impliqués dans le phénomène étudié.
Mais il n’est pas certain, même en isolant le plus grand nombre de mutants possibles,
que le crible de mutants choisi ait permis de toucher tous les gènes impliqués
dans le phénomène étudié :
– les mutations de certains gènes peuvent être létales et ne peuvent être sélectionnées;
– certains gènes peuvent être redondants, de sorte que la mutation d’un d’entre eux
sera sans effet sur le phénotype qui demeure sauvage;
– le crible de mutants utilisé est inefficace pour isoler des mutants
de certains des gènes impliqués dans le caractère ou le phénomène, particulièrement
quand il s’agit de gènes régulateurs impliqués dans des cascades d’expression
(par exemple gènes du développement, ou du cycle cellulaire) en raison du type
d’interaction de leur produit avec les gènes (interaction protéine-ADN) ou les
produits des gènes (interaction protéine-protéine) dont on peut isoler des mutants.
C’est pourquoi les généticiens ont assez vite découvert l’intérêt qu’il y avait de
compléter leur analyse génétique des mutants « directs », obtenus par un crible défini,
par l’analyse de « révertants ».
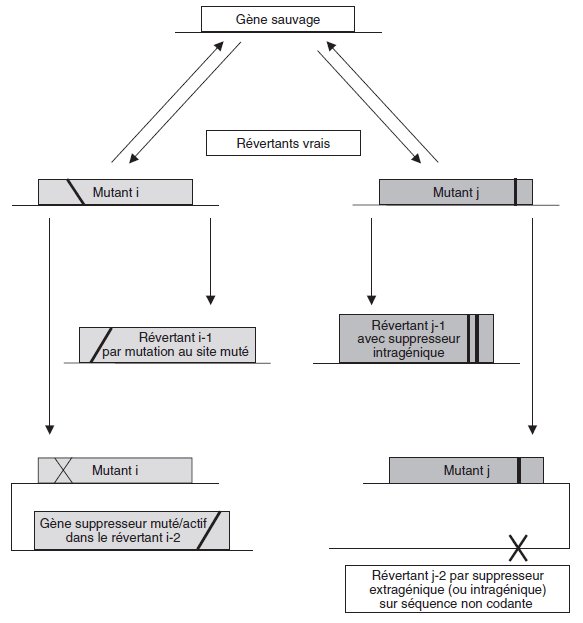
Définition : un « révertant » est un mutant de mutant direct qui a recouvré le phénotype
originel dont était issu le mutant direct (en général le phénotype sauvage si le
mutant est issu d’une SSR).
L’analyse des révertants va se révéler très informative car
– elle permet d’identifier des gènes non identifiés ou non identifiables par la mutagenèse
directe;
– elle apporte des précisions fonctionnelles, voire moléculaires, sur la nature des
différentes mutations affectant les gènes identifiés chez les mutants directs;
– elle permet de mettre en évidence des interactions fonctionnelles entre gènes.
Remarque 1. Les révertants sont, sauf exceptions, obtenus par un crible de
sélection inverse de celui qui a permis d’avoir le mutant direct.
À partir d’un
mutant direct de SSR, sélectionné par un « crible négatif », on
obtiendra un révertant ayant recouvré le phénotype sauvage, par un « crible
positif ».
À l’inverse les révertants issus de mutants directs obtenus par crible
positif, seront sélectionnés à travers un crible négatif.
Remarque 2. Une souche originelle SSR a un génotype et un phénotype sauvages
et les mutants directs ont un phénotype muté parce qu’ils ont un génotype
muté.
Par contre, si les révertants ont recouvré un phénotype sauvage, rien ne permet de
dire que leur génotype est sauvage, c’est pourquoi on les désigne souvent comme des
« révertants phénotypiques ».
L’analyse génétique, éventuellement moléculaire, doit
permettre de distinguer les révertants « vrais » qui ont recouvré un génotype sauvage,
et les autres qui présentent en fait un génotype muté, soit qu’une deuxième mutation
ait remplacé la première (révertants « au site muté »), sans avoir l’effet phénotypique
de celle-ci, soit qu’une deuxième mutation, en un autre site (dans le même
gène ou hors de ce gène) ait la propriété de « supprimer » l’effet de la mutation
directe, ce qui rétablit le phénotype sauvage.
Dans ce dernier cas la mutation est appelée « mutation suppresseur » ou mutation
à effet suppresseur.
Le suppresseur est intragénique si la mutation touche, en un
autre site, le même gène que la mutation directe; il est dit « extragénique » s’il est
extérieur au gène porteur de la mutation directe.
Un suppresseur extra-génique peut toucher un gène qui sera alors appelé « gène
suppresseur » mais il peut aussi toucher une séquence non exprimée mais signifiante
dans l’expression du génome, notamment celle du gène porteur de la mutation
directe (figure 7.1).
Remarque sémantique et conceptuelle.
La figure 7.1 montre bien qu’une mutation
suppresseur ne supprime pas, au sens strict, la mutation originelle ou
directe; il est correct de dire que c’est l’effet de la mutation suppresseur qui
supprime l’effet (ou le non effet) de la mutation directe.
Par contre la mutation reverse au site muté supprime bien, au sens strict, la mutation
directe et par là l’effet (ou le non effet) de cette mutation en lui substituant son
effet propre.
Analyse
génétique formelle des révertants :
A - Taux de réversion
:
La fréquence de révertants est toujours, sauf exceptions, plus faible que la fréquence
de mutants directs, de l’ordre de 10 à 1 000 fois plus faible, car la fréquence de
mutation d’un phénotype donné est une fonction de la taille de la « cible » qui doit
être mutée.
Au départ, il est d’autant plus facile d’avoir des mutants directs que le nombre de
gènes impliqués dans le phénotype étudié est grand, puisqu’il suffit a priori d’en
toucher un seul pour avoir un phénotype mutant.
Pour les mutants directs la taille de
la « cible » correspond physiquement à la longueur totale des séquences mutables,
notamment si le phénotype muté dépend de mutations de perte de fonction.
Pour ce qui est des révertants, la taille de la cible est obligatoirement plus faible
puisque la mutation doit toucher un site particulier, celui qui, dans le gène ou hors de
lui, aura un effet suppresseur.
Le fait que certains mutants d’un gène donne assez facilement des révertants alors
que d’autres mutants du même gène n’en donnent que très rarement, voire jamais, ne
pourra donc être interprété que comme une conséquence de la nature différente des
mutations directes affectant le gène chez les différents mutants.
Ces faits seront discutés
et interprétés plus loin, mais montrent déjà l’intérêt des révertants dans l’approfondissement
de l’analyse des mutants directs.
B - Mise en évidence d’une mutation suppresseur chez un révertant
:
L’analyse génétique d’un révertant consiste en premier lieu à savoir si on peut mettre
en évidence une mutation suppresseur, et donc le maintien de la mutation directe
(originelle).
Pour ce faire, on croise le révertant avec la souche d’origine du mutant direct,
c’est-à-dire la SSR dans le cas où le mutant direct en est issu, puis on observe les produits de la méiose afin de mettre en évidence une éventuelle recombinaison entre
la mutation directe, si elle a été maintenue, et le suppresseur, s’il existe et qu’il n’est
pas lié trop fortement à la mutation directe.
Dans ce cas on voit réapparaître le
phénotype mutant dans la descendance du croisement et le révertant est dit de
seconde classe (voir tableau 7.1).
Figure 7.1 Filiation et variété des mutants directs et de leurs révertants respectifs. Deux mutants i et j issus d’un même génotype sauvage ne seront mutés ni au même
site, ni de la même façon (par exemple l’un est non-sens et l’autre est un mutant de
décalage du cadre de lecture); les révertants issus d’un même mutant peuvent avoir
des génotypes très différents, même s’ils ont le même phénotype (ce sont des
révertants !).
Dans le cas contraire, quand aucun descendant ne présente de phénotype muté,
soit qu’il n’y a pas eu de recombinaison entre la mutation directe et le suppresseur,
car celui-ci est génétiquement très lié, notamment en cas de suppresseur intragénique,
soit que la mutation directe a disparu au profit d’une deuxième mutation au
site, les révertants sont dits de première classe (voir tableau 7.1).
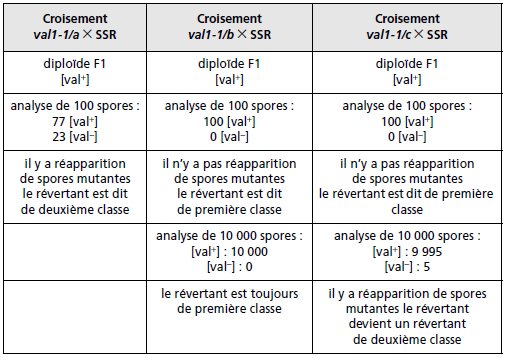
Exemple de croisements permettant de statuer
sur la présence d’un suppresseur chez des révertants
:
On a obtenu à partir d’une souche haploïde de levure, prototrophe pour la
valine, des mutants auxotrophes dans un gène désigné par VAL1.
À partir
d’un de ces mutants simples nommé val1-1, on obtient trois révertants
notés val1-1/a, val1-1/b et val1-1/c, qu’on croise avec la SSR (tableau 7.1).
a) Les révertants de seconde classe et leur interprétation génétique
La méiose du diploïde issu du croisement entre le SSR et val1-1/a donne
23 spores [val–] (tableau 7.1, colonne 1).
La réapparition du phénotype
muté caractérise le « révertant de deuxième classe » dont l’interprétation
fonctionnelle est sans ambiguïté : c’est un révertant avec suppresseur,
puisque la réapparition du phénotype muté atteste que la mutation directe
n’avait pas disparu, que son effet était supprimé chez le révertant par celui
d’une deuxième mutation dite suppresseur, ces deux mutations ayant été
séparées par recombinaison génétique lors de la méiose chez le diploïde,
ce qui conduit notamment à la reconstitution d’un génome mutant direct.
TABLEAU 7.1 ANALYSE GÉNÉTIQUE DE RÉVERTANTS PAR CROISEMENT AVEC LA SSR.
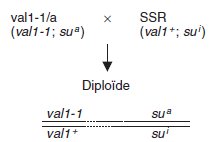
En effet on peut écrire les croisements, sur le plan génotypique, de la façon
suivante, où val1+ et val1-1 désignent les allèles du gène VAL1, et sua la
mutation suppresseur (su pour suppresseur et a pour actif), son homologue
sauvage étant désigné par sui (su pour suppresseur et i pour inactif) :
Compte tenu de la recombinaison génétique possible (par assortiment
aléatoire des chromosomes si il y a indépendance physique, par crossingover
si il y a liaison) on peut attendre quatre types possibles de spores, dont
on sait à l’avance les génotypes et, pour trois d’entre elles, les phénotypes :
parental 1 : (val1-1; sua) de phénotype [val+]
parental 2 : (val1+; sui) de phénotype [val+]
recombiné 1 : (val1-1; sui) de phénotype [val–]
recombiné 2 : (val1+; sua) de phénotype [val?]
Les spores [val–] ne peuvent qu’être issues d’une recombinaison génétique
entre la mutation directe et son suppresseur et attestent de l’existence de
celui-ci.
Que peut-on dire du phénotype associé au génotype (val1+; sua) qui n’a
jamais été encore rencontré ?
– A priori rien d’un point de vue physiologique, même si on peut supposer
que l’allèle sua a un effet spécifique sur celui de l’allèle val1-1 et qu’il
ne devrait pas entraver l’effet dominant de l’allèle val1+, ce qui aboutirait
dans ce cas au phénotype [val+].
– D’un point de vue ségrégatif, on peut remarquer qu’il y a 3/4 de spores
[val+] et 1/4 de spores [val–], ce qui correspondrait très exactement au
résultat attendu si on a un gène suppresseur indépendant du gène VAL1
et que le génotype (val1+; sua) est de phénotype [val+].
Mais on pourrait
aussi imaginer que le génotype (val1+; sua) est [val–] et que les deux
gènes sont assez liés pour que la fréquence des spores recombinées ne
soit que de 25 % au total (distance de 12,5 u.r.) !
Des expérimentations supplémentaires sont nécessaires pour résoudre
cette question sans ambiguïté, comme par exemple une analyse de tétrades,
possible chez la levure.
– L’observation de tétrades avec quatre spores [val–] ne pourrait être interprétée
que comme un ditype recombiné et la preuve que le génotype
(val1+; sua) est [val–].
Dans ce cas les DP contiendraient quatre spores
prototrophes et les T, deux auxotrophes et deux prototrophes.
– L’observation de tétrades avec trois spores [val+] et une seule spore [val–]
ne pourrait être interprétée que comme un tétratype et la preuve que le
phénotype (val1+; sua) est [val+].
Dans ce cas les DP contiendraient
quatre spores prototrophes et les DR, deux auxotrophes et deux prototrophes
(construire les tableaux d’analyse de tétrades à titre d’exercice).
b) Les révertants de première classe et leur interprétation génétique
À la méiose, chez les diploïdes issus des croisements entre la SSR et les
révertants val1-1/b ou val1-1/c (tableau 7-1, colonnes 2 ou 3), on n’observe
dans un premier temps, sur 100 spores testées, que des spores haploïdes de
phénotype [val+].
Ce résultat, caractérisé par l’absence de réapparition du
phénotype muté, définit le « révertant de première classe », dont l’interprétation
fonctionnelle est ambiguë.
Ce peut être un révertant vrai, de génotype sauvage, ou un révertant par
mutation au site muté, de génotype non sauvage mais fonctionnel, ou un
révertant porteur d’une mutation suppresseur très proche du site de la mutation
directe (suppresseur intragénique ou dans un gène contigu) de sorte
que la recombinaison (par CO) est trop rare pour être mise en évidence par
le nombre de méioses étudiées.
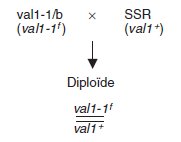
Dans le cas d’un révertant par mutation au site muté, si val1+ et val1-1f
désignent respectivement l’allèle sauvage et l’allèle révertant muté au site,
le croisement peut s’écrire :
ce qui donne, par ségrégation 2/2, deux types de spores de même phénotype
[val+].
On peut tenter de lever l’ambiguïté d’interprétation pour les révertants de
première classe en accroissant le nombre de méioses étudiées afin, si un
suppresseur très lié existe, d’augmenter la probabilité d’observer au moins
un gamète, une spore, recombiné de génotype mutant. Dans l’exemple du
révertant val1-1/c, l’étude de 10 000 spores, soit environ 2 500 méioses a
permis de faire apparaître cinq spores [val–].
L’observation d’une seule spore mutée (à condition que sa fréquence soit supérieure au taux de mutation
!) suffit à lever l’ambiguïté et à considérer que val1-1/c est un révertant
de deuxième classe avec un suppresseur très lié à la mutation directe.
Remarque 1. L’intensité de la liaison génétique ne permet nullement de considérer
que le suppresseur de val1-1/c est intra-génique, mais elle justifie de se
poser la question et de trouver le moyen expérimental de la résoudre.
Remarque 2. Les suppresseurs présents chez val1-1/a et val1-1/c ne sont pas
les mêmes car ils sont indépendants, ils doivent être distingués dans l’écriture
et seront respectivement notés suaa et suca.
Remarque 3. Le calcul de la distance génétique entre la mutation directe et le
suppresseur suca est limité par la méconnaissance du phénotype des spores
(val1+; suca).
Si ce phénotype est [val+], la fréquence totale des spores recombinées
est égale à 10/10 000, car il faut rajouter en moyenne 5 spores [val+]
aux 5 spores [val–].
Si ce phénotype est [val–], la fréquence totale des spores
recombinées est égale à 5/10 000, car toutes les spores recombinées ont alors
le même phénotype.
Remarque 4. On peut imaginer, chez un révertant par deuxième mutation au
site de la mutation directe, qu’un crossing-over intra-codon, exceptionnel par
sa rareté, puisse survenir au site muté val1-1f et reconstitue un site muté
direct, si la paire de bases mutée chez le révertant n’est pas la même que la
paire de base mutée chez le mutant direct val1-1 (exemple : codon sauvage
GAG, mutation stop TAG, codon révertant TAC : un crossing-over entre GAG
et TAC donne deux allèles GAC et TAG et reconstitue le mutant STOP val1-1
et un nouveau mutant GAC, le crossing-over intra-codon peut donc être mutationnel).
En résumé, le but du croisement d’un révertant de phénotype sauvage avec la SSR, de phénotype et de génotype sauvage, consiste à construire un diploïde (F1 si
on est chez un diplobiontique) au génotype double hétérozygote pour la mutation
directe et l’éventuel suppresseur, puis à tester l’éventuel retour dans la descendance
(spores haploïdes ou F2) de phénotypes mutants attestant que la mutation originelle
n’avait pas disparu et que son effet était supprimé, chez le révertant, par celui d’une
mutation suppresseur, ces deux mutations ayant été dissociées par recombinaison
génétique lors de la méiose.
Si tel est le cas le révertant est dit de « deuxième classe ».
Les révertants dits de « première classe » ne présentent pas, après croisements avec
la SSR de descendants de phénotype muté.
Ce résultat peut admettre plusieurs interprétations
(révertant vrai, révertant au site muté, ou suppresseur génétiquement très
lié à la mutation directe).
La répétition des croisements peut éventuellement faire passer le statut d’un
révertant de la première classe à la seconde.
NB : en génétique bactérienne, pour des raisons spécifiques aux méthodes de
« croisements », l’étude des révertants sera réalisée à travers des croisements avec la
souche mutante et non la souche sauvage (voir exercices partie 2).