



















Physiologie nasale :
A – INTRODUCTION :
La physiologie des fosses nasales recouvre un ensemble de phénomènes complexes dont les mécanismes ne sont pas encore totalement connus.
Les principales fonctions sont la ventilation, la défense des voies aériennes supérieures et l’olfaction.
 Nous abordons les principales fonctions de la muqueuse nasosinusienne, en intégrant les données les plus récentes sur la ventilation nasale, le rôle de la muqueuse nasale et sinusienne dans la réponse inflammatoire à des agressions spécifiques (allergie, infection) ou non (pollution) et le système immunitaire attaché à la muqueuse nasosinusienne (NALT) qui est également un sujet de recherche pour des thérapeutiques futures (vaccination) et pour la compréhension de la physiopathologie d’affections courantes telles les rhinosinusites allergiques ou la polypose.
Nous abordons les principales fonctions de la muqueuse nasosinusienne, en intégrant les données les plus récentes sur la ventilation nasale, le rôle de la muqueuse nasale et sinusienne dans la réponse inflammatoire à des agressions spécifiques (allergie, infection) ou non (pollution) et le système immunitaire attaché à la muqueuse nasosinusienne (NALT) qui est également un sujet de recherche pour des thérapeutiques futures (vaccination) et pour la compréhension de la physiopathologie d’affections courantes telles les rhinosinusites allergiques ou la polypose.
La physiologie nasosinusienne est complexe et son étude, en constant progrès, fait l’objet de nouvelles découvertes dont les applications cliniques sont nombreuses en pratique quotidienne : antihistaminique, corticoïdes, antileucotriènes, etc.
B – RAPPEL ANATOMIQUE ET HISTOLOGIQUE :
1- Anatomie macroscopique :
Les cavités nasales sont limitées en bas par les processus palatins des maxillaires, en haut et en avant par les cartilages alaires, triangulaires, puis les os du nez.
En arrière, l’os ethmoïdal forme la voûte nasale.
L’orifice antérieur ou vestibule est un canal, avec un revêtement cutané en avant et muqueux en arrière.
Les valves nasale et septoturbinale font partie du vestibule.
L’orifice postérieur ou choane comprend une partie supérieure, l’arcade choanale, et une inférieure, le seuil choanal.
Le récessus ethmoïdosphénoïdal surplombe l’orifice choanal.
Le plancher des cavités nasales, en forme de gouttière, est limité médialement par le septum et latéralement par la paroi latérale ; celle-ci, ou paroi turbinale, comprend le cornet inférieur, les cornets ethmoïdaux avec leurs méats respectifs : c’est la paroi de communication avec les cavités sinusiennes.
La voûte nasale comprend essentiellement la fente olfactive.
2- Histologie :
La muqueuse nasale comprend une muqueuse respiratoire et une muqueuse olfactive. Seule la muqueuse respiratoire est envisagée dans cet article.
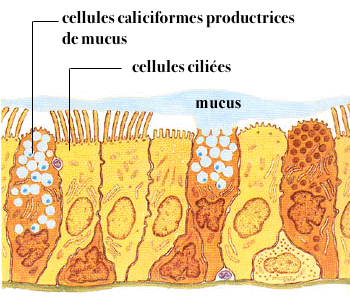
La muqueuse respiratoire comprend un épithélium de surface composé d’une assise unistratifiée de cellules prismatiques ciliées de type respiratoire qui possèdent environ 200 cils de 5 à 10 µm de long.
En plus de ces cellules, on observe des cellules caliciformes mucipares, des cellules de remplacement, profondes, donnant un aspect pluristratifié.
Entre les cellules épithéliales émerge par endroit l’ostium des glandes tubuloacineuses qui se trouvent dans le chorion.
On y trouve également des cellules mélaniques, des cellules du système endocrinien diffus, des lymphocytes.
Le chorion est classiquement décrit avec trois réseaux :
– le réseau superficiel comprend du fibrocollagène lâche, renfermant un riche réseau vasculaire capillaire et nerveux ;
– le réseau moyen est surtout marqué au niveau du cornet inférieur ; il comprend des artérioles perpendiculaires au plan muqueux qui anastomosent les réseaux profonds et superficiels capillaires ;
– le réseau fibrocollagénique profond, dense, adhérant au périoste, comprend un réseau artérioveineux anastomosé en nappe.
C – VENTILATION NASALE :
Le passage de l’air inspiré et expiré dans les cavités nasales a été décrit avec précision par les travaux de Swift et Proctor à partir de pièces anatomiques.
L’essentiel du passage de l’air inspiré se fait entre le cornet inférieur et le cornet moyen.
Le plancher nasal et le toit sont les régions les moins ventilées.
Il est classique de signaler que la taille et la direction des orifices narinaires peuvent influencer la direction et la vitesse du flux aérien, mais ce paramètre n’a pas été à ce jour étudié scientifiquement.
À l’expiration, la répartition est plus dispersée, en particulier avec une bonne distribution vers les régions olfactives.
Ces résultats ne tiennent toutefois pas compte des modifications induites par les modifications des cartilages et des muscles de la cavité nasale, dont le rôle in vivo demeure méconnu.
On prête à Kayser, dès 1895, la première description scientifique des modifications alternatives de la perméabilité nasale dénommée « cycle nasal ».
Depuis cette première description, de nombreux travaux ont été réalisés pour comprendre ce phénomène et ses mécanismes.
L’analyse de ces différents travaux permet de constater que seuls 70 à 80 % de la population adulte présentent ces modifications cycliques.
Le cycle nasal est caractérisé par une alternance de vasoconstriction et de vasodilatation qui touche tout le tissu vasculaire, dont les principales localisations sont le cornet inférieur et le septum nasal.
Sa périodicité peut varier de 1 à 5 heures, mais on peut également observer des modifications vasculaires asymétriques et non rythmiques.
Le cycle nasal touche également la muqueuse des cavités sinusiennes maxillaires et ethmoïdales.
Le cycle nasal se modifie avec l’âge, une étude sur 361 patients, considérés comme indemnes de pathologies rhinosinusiennes, suggère que des modifications du cycle nasal peuvent survenir jusqu’à l’âge de 16 ans en raison des modifications anatomiques et qu’ensuite une certaine stabilité est observée.
Le mécanisme régissant ces modifications vasomotrices a été récemment rappelé par Lung.
Le réseau vasculaire de la muqueuse nasale comprend des vaisseaux de résistance (artères et artérioles) et des vaisseaux de capacitance (plexus caverneux et veines).
Il est classique de décrire, au niveau du cornet inférieur, trois circuits vasculaires superposés :
– le plus profond correspond au système résistif formé par des anastomoses artérioveineuses siégeant dans le chorion ; un sphincter musculaire lisse permet de réguler les passages directs entre systèmes artériel et veineux ;
– le circuit intermédiaire est formé par des plexus caverneux, éléments capacitaires du système ; ces structures avalvulaires résident dans le chorion moyen ; leur volume serait sous la dépendance des anastomoses artérioveineuses en amont et de sphincters veineux en aval ;
– le plus superficiel comprend les terminaisons des rameaux ascendants issus du réseau artériel sous-périosté et/ou souspérichondral ; ces artérioles donnent naissance à un réseau capillaire de type fenêtré qui se distribue sous la membrane basale aux structures épithéliales et glandulaires ; cette structure est un lieu d’échange, essentiel pour la défense et l’information de l’organisme.
Au total, on constate que chaque système peut être régulé indépendamment, mais également interagir sur les autres systèmes vasculaires, créant les conditions d’une indépendance au sein d’un système d’interdépendance.
La régulation de cette circulation est également complexe.
Les vaisseaux de la muqueuse nasale reçoivent des stimulations constantes par le système sympathique, alors que le système parasympathique aurait une activité plus irrégulière.
Les fibres sympathiques préganglionnaires proviennent des neurones situés dans les cornes antérieures de la moelle thoracique, entre C6 et D2.
Certaines se terminent dans les ganglions cervicaux inférieur et moyen, mais la plupart atteignent le ganglion cervical, pour faire synapse.
Les fibres postsynaptiques se dirigent, au travers du nerf grand pétreux, vers le ganglion sphénopalatin.
Quelques fibres accompagnent la carotide interne puis la vascularisation à destinée nasale.
Les fibres sympathiques contiennent soit de la noradrénaline (NAR), soit de la noradrénaline et du neuropeptide Y (NPY) qui sont tous deux des médiateurs vasoconstricteurs.
Des récepteurs alpha1- ou alpha2-adrénergiques sont présents sur la muqueuse nasosinusienne.
Les fibres nerveuses sympathiques stimulent, par la libération de noradrénaline, les récepteurs alpha1 et alpha2 dont la distribution est mixte pour les vaisseaux de capacitance et presque exclusive (alpha2) pour les vaisseaux de résistance.
La stimulation sympathique est à l’origine d’une vasoconstriction du système résistif par un mécanisme alpha-adrénergique et d’une vasoconstriction des vaisseaux de capacitance par le même mécanisme, mais également par une stimulation non adrénergique et non cholinergique.
Une étude récente in vitro sur la muqueuse du cornet moyen a permis de confirmer que la somatostatine et le neuropeptide Y provoquent une vasoconstriction selon un mécanisme non adrénergique-dépendant, peut-être par stimulation de récepteurs spécifiques.
Cette étude in vitro suggère également un rôle modulateur de ces médiateurs sur l’action vasoconstrictrice de la noradrénaline.
D’autre part, des récepteurs à l’histamine ont été récemment identifiés dans l’endothélium des vaisseaux du cornet inférieur. Enfin, l’étude sur l’animal a montré que la régulation de la libération de NAR et NPY serait sous la dépendance d’un récepteur adrénergique alpha-préjonctionnel.
Toutes ces données confirment la complexité des mécanismes régulant la vasomotricité nasale.
À l’état pathologique, de nombreux autres médiateurs viennent troubler cette délicate homéostasie (histamine, leucotriènes, etc).
Les fibres parasympathiques proviennent du noyau salivaire supérieur (muco-lacrymo-nasal) puis se dirigent vers le ganglion géniculé et se rendent, par le nerf grand pétreux, vers le ganglion ptérygopalatin où elles font relais pour donner des fibres à destinées nasale et lacrymale ; des fibres postganglionnaires innervent également des glandes et quelques vaisseaux.
La stimulation des fibres parasympathiques libère de l’acétylcholine, du vaso intestinal peptide (VIP) et peut-être d’autres neuropeptides.
Cinq types de récepteurs muscariniques (m1 à m5) ont été décrits dans des études in vitro ou in vivo.
In vivo, le rôle respectif de chacun demeure encore l’objet de controverses.
La stimulation du système parasympathique entraîne surtout une augmentation de la sécrétion de mucus.
La présence de VIP est signalée dans les polypes et le cornet inférieur dans l’épithélium et les glandes sous-muqueuses. Plusieurs facteurs peuvent modifier le rythme et l’amplitude du cycle nasal.
Récemment, Ishii a étudié les modifications du cycle nasal lors d’atteintes spécifiques du système nerveux autonome.
Ainsi, chez les patients atteints d’une paralysie faciale où des anomalies du système parasympathique sont signalées, peu d’anomalies du cycle nasal sont observées.
Lors d’atteintes du système sympathique (syndrome de Horner), aucune perturbation du cycle nasal n’est observée.
Ces deux constatations confirment la probable régulation centrale des modifications vasculaires cycliques de la muqueuse nasale et sinusienne.
Le centre régulant ce rythme nasal est peut-être localisé dans le noyau suprachiasmatique de l’hypothalamus.
Les résistances nasales varient également selon la position de la tête ou du sujet (assis, décubitus, etc).
Des pressions, appliquées sur certaines zones du corps, peuvent modifier les résistances nasales.
Ainsi, lors du décubitus latéral, des modifications sont observées alternativement en fonction des pressions appliquées sur la surface cutanée.
La consommation importante de tabac augmente les résistances nasales mesurées par rhinomanométrie antérieure.
La sensation de confort nasal demeure une donnée subjective dont la mesure n’est pas évaluable actuellement, même avec la rhinomanométrie.
Cette sensation est d’ailleurs très subjective puisqu’elle peut être corrélée à des paramètres culturels.
La sensation la plus importante du passage de l’air est obtenue dans la partie antérieure de la cavité nasale.
Les mesures de résistance de l’air dans la filière nasale à l’aide d’un pléthysmographe chez 45 sujets normaux adultes confirment que la zone de résistance se situe dans les 4 premiers centimètres pour une fosse nasale non décongestionnée.
Cette région se réduit aux 2 premiers centimètres dans une fosse nasale décongestionnée.
Ces résultats expliquent l’absence actuelle de consensus sur la localisation exacte de la région dite de « la valve nasale ».
Les études concernant les mécanismes régissant cette sensation de confort nasal amènent à des conclusions contradictoires.
Ainsi, Eccles a montré que l’inhalation de menthol améliore la sensation de confort nasal sans aucune modification des résistances nasales.
Le L-menthol modifierait directement le transport calcique dans les fibres nerveuses.
Naito rapporte également un phénomène similaire après stimulation par le menthol du nerf grand palatin.
Une stimulation des récepteurs au froid présents dans la muqueuse nasale serait à l’origine de ce phénomène.
De ce fait, il semble plus intéressant de mesurer le coefficient d’accélération de l’air inspiré pour obtenir une meilleure corrélation avec la sensation subjective de confort nasal.
L’analyse de la sensation du passage de l’air a également été étudiée après la pulvérisation de xylocaïne ou de sérum dans les cavités nasales chez 50 patients.
Cette étude, faite en double aveugle avec une analyse par rhinomanométrie et échelle visuelle analogique, ne met pas en évidence de différence entre les deux groupes.
Cette constatation minimiserait le rôle des récepteurs sensitifs dans la perception du flux aérien.
De même, Aldren a montré que l’application par crème d’un anesthésique chez 25 sujets modifie le confort nasal sans aucune modification objective des résistances nasales.
D’autre part, Cauna, pour expliquer la sensation de confort nasal, ne trouvait, par une étude histologique, qu’un seul type de récepteur correspondant à la terminaison de fibres amyélinisées cholinergiques auxquelles il attribuait un rôle prépondérant pour expliquer cette sensation.
Néanmoins, d’autres récepteurs ont vraisemblablement un rôle dans cette perception de confort nasal puisque, jusqu’à présent, aucune publication n’a relaté la survenue d’une sensation d’obstruction nasale après la destruction chimique ou chirurgicale de la branche maxillaire du nerf trijumeau.
D’autres auteurs ont d’ailleurs mis en évidence des récepteurs thermiques au chaud et au froid dans le vestibule nasal antérieur (cutané) et postérieur (muqueuse).
La prépondérance de la respiration par voie nasale est encore mal connue et fait l’objet de débats contradictoires.
Pour Kluemper, par exemple, l’obstruction nasale chez l’enfant peut être à l’origine de dysfonction buccolinguale, alors que pour d’autres aucune corrélation spécifique entre un type de malocclusion et une déformation faciale n’est confirmée scientifiquement.
Récemment, Laine-Alava et al ont montré, avec une étude sur 361 sujets, que l’équilibre entre respiration buccale et nasale se modifiait jusqu’à l’âge de 16 ans et que les résistances nasales étaient plus importantes pendant l’inspiration, peut-être en raison de l’activité des muscles de la région des alaires.
Une autre étude récente a montré qu’une obstruction nasale bilatérale provoquée lors du sommeil avait pour conséquence une diminution de la saturation en oxygène du sang périphérique, suggérant un rôle prépondérant de la ventilation nasale pendant le sommeil.
Finalement, l’analyse de ces différents travaux confirme que les mécanismes neurophysiologiques soutenant la transformation du stimulus ventilatoire en une sensation de confort demeurent spéculatifs et inconnus.
D – HUMIDIFICATION ET RÉCHAUFFEMENT DES CAVITÉS NASALES :
La notion de confort nasal est également liée à la sensation d’humidification des cavités nasales.
Elle résulte du contact et des échanges entre l’air inspiré et le mucus. Le réseau vasculaire sousépithélial fenêtré jouerait un rôle essentiel dans cet équilibre, ainsi que les glandes séreuses.
Cette humidification est essentielle à la protection de la muqueuse en maintenant les caractéristiques rhéologiques du mucus formé à 95 % d’eau.
Les mécanismes la régulant restent mal connus, car difficiles à étudier.
Le rôle de la ventilation nasale dans le réchauffement de l’air inspiré est important ; ce système peut fonctionner et s’adapter à des conditions extrêmes (de – 10° à + 40° ; haute altitude, effort extrême) ; sa régulation et son mécanisme sont encore mal connus ; la richesse artérioveineuse joue vraisemblablement un rôle majeur dans cette adaptation, la surface de la muqueuse des cornets est d’ailleurs très importante chez les animaux vivant dans des conditions climatiques extrêmes.
Il n’y a pas néanmoins d’étude chez l’homme qui confirme des modifications objectives de la vascularisation turbinale en cas de modifications rapides des conditions atmosphériques (température, humidité).
E – SÉCRÉTIONS NASALES :
Les sécrétions nasales sont formées essentiellement par les glandes nasales (glycoprotéines du mucus), l’exsudation plasmatique, les larmes et les phénomènes de condensation de vapeur d’eau.
Les principales glandes nasales sont les glandes à cellules caliciformes et les glandes séreuses.
La cytologie des sécrétions nasales a été surtout étudiée lors de pathologies inflammatoires comme la rhinite à éosinophiles ou lors d’infections.
L’analyse électrophorétique des sécrétions nasales, en particulier des protéines chez l’adulte, révèle qu’il n’y a pas de différence selon le sexe ou l’âge du patient et que la répartition se fait selon un gradient de poids moléculaire allant de 14 à 70 kDa.
Des immunoglobulines sont également présentes (IgA, IgM, IgG, IgE) ainsi que de nombreuses enzymes (endopeptidases, antileucoprotéases, aminopeptidases) et peptides (substance P, CGRP [calcitonin gene related peptide]).
Les fonctions de ces sécrétions sont multiples : antioxydante, humidification, adhésion et élimination de micro-organismes ou de particules, etc.
La régulation du volume et de la rhéologie des sécrétions nasales est complexe et encore incomplètement connue.
L’activité sécrétoire de la muqueuse nasale est également sous contrôle du système neurovégétatif.
La stimulation des fibres parasympathiques augmente les secrétions nasales, en particulier par une action directe sur les glandes séreuses.
De nombreux peptides (substance P, VIP [vaso intestinal peptide], CGRP, GRP [gastrine releasing peptide]) sont capables in vitro de stimuler directement les sécrétions glandulaires.
Le GRP, par exemple, est signalé dans les nerfs se distribuant aux glandes sous-muqueuses qui possèdent des récepteurs spécifiques pour ce médiateur.
Il stimulerait ainsi la sécrétion de mucoglycoprotéines.
Le système veineux participerait également, par l’exsudat plasmatique qu’il produit pendant les modifications cycliques de vasodilatation et vasoconstriction, aux modifications sécrétoires observées lors d’infection.
Ces veines ayant un endothélium fenêtré, l’augmentation de la fréquence du cycle s’accompagnerait d’une augmentation parallèle du volume des sécrétions et, ainsi, participerait également de façon indirecte au système de défense.
La stimulation unilatérale par un air sec déclenche une rhinorrhée bilatérale chez la plupart des sujets.
Cette stimulation est peut-être médiée par les terminaisons sensitives, puisque l’application d’un anesthésique local du côté de la stimulation inhibe la sécrétion des deux côtés, alors qu’elle ne modifie pas la stimulation par la métacholine.
Les composants anatomiques et les voies associatives de l’arc réflexe à l’origine de cette sécrétion induite par des modifications des propriétés physiques de l’air restent à définir.
La localisation et le rôle des récepteurs bêta-adrénergiques restent encore à évaluer.
D’après Woodhead, le plus grand nombre est de type bêta-2 et se localise dans les canaux glandulaires, suggérant un rôle potentiel dans la composition électrolytique des secrétions nasales.
F – INNERVATION NASOSINUSIENNE :
1- Généralités :
Les nerfs sensitifs sont des nerfs issus du trijumeau qui se distribuent par les nerfs ethmoïdaux et nasaux postérieurs qui innervent les vaisseaux, les glandes et l’épithélium.
Les nerfs nociceptifs sont des nerfs non myélinisés de type C qui ne possèdent pas d’organes terminaux spécialisés comme ceux présents dans le tissu cutané.
Néanmoins, des formations plexiformes ont été décrites aux extrémités de fibres non myélinisées sensitives dans la lamina propria et entre les cellules épithéliales.
Deux types de fibres conduisent l’influx nociceptif : les fibres A-ä conduisent la douleur aiguë.
Les fibres C seraient plutôt impliquées dans la réponse à une stimulation plus complexe et dans les douleurs chroniques.
Ces neurones chimiosensibles et mécanothermiques peuvent être stimulés par des médiateurs inflammatoires : histamine, bradykinine, sérotonine, mais également par des modifications ioniques ; ion K+, ion H+.
La stimulation des nerfs sensitifs est à l’origine de mécanismes réflexes se traduisant par une toux, des éternuements et une hypersécrétion.
Les prostaglandines et les leucotriènes modifient le niveau de dépolarisation de ces neurones en favorisant leur stimulation.
D’autres substances peuvent stimuler ces nerfs : cigarette, capsaïcine, SO2 (dioxyde de soufre).
Après stimulation des fibres de type C, on peut observer un réflexe parasympathique d’origine centrale se traduisant par une vasodilatation.
Il existe également une dépolarisation de proche en proche rétrograde, à l’origine d’une libération de neuromédiateurs (réflexe d’axone), amplifiant la vasodilatation et la perméabilité vasculaire localement.
Lors de la dépolarisation du nerf, une combinaison variable en nature et en dose des neuropeptides est délivrée au contact des glandes et des vaisseaux.
2- Différents neuropeptides :
Les tachykinines sont des peptides dont font partie la substance P (SP), les neurokinines A (NKA) et B (NKB).
Trois types de récepteurs sont individualisés, respectivement NK1 pour SP, NK2 pour NKA et NK3 pour NKB.
* Substance P :
La substance P induit une vasodilatation, une sécrétion de mucus et une exsudation plasmatique ; elle a été essentiellement signalée autour des artères, mais également un peu à proximité du système veineux, les cellules glandulaires et épithéliales nasales.
Récemment, l’identification des gènes codant pour les récepteurs NK1, NK2, NK3 a permis à Shirasaki de localiser spécifiquement sur le cornet inférieur les récepteurs NK1, principalement dans l’épithélium et les glandes sous-muqueuses, avec une moindre répartition vers les vaisseaux.
In vitro, la substance P déclenche une sécrétion de lactoferrine (glande séreuse) et de glycoprotéine (glandes muqueuses).
In vivo, en revanche, la stimulation par la substance P ne modifie pas les secrétions nasales, peut-être en raison d’une destruction rapide in situ par les endopeptidases (NEP).
D’autre part, la substance P peut augmenter l’expression de cytokines tel IL1 (interleukine 1) et IL6 chez le sujet non allergique ou tel IL1, IL2, IL3, IL4, IL5, IL6, TNFalpha (tumour necrosis factor) chez le sujet allergique, confirmant la plus grande réactivité des sujets allergiques à la substance P.
Le rôle physiologique de la substance P reste néanmoins encore à préciser.
Baraniuk, par exemple, fait remarquer que si, après stimulation antigénique, son dosage est élevé dans les sécrétions et s’associe à une augmentation des éosinophiles dans la muqueuse, son élévation dans la bronchite chronique ne s’accompagne d’aucune éosinophilie tissulaire.
Bascom, étudiant des sujets présentant une réaction de congestion nasale au contact de la fumée de tabac, a trouvé que ses patients avaient une réactivité à la substance P supérieure à celle d’un groupe-témoin.
* CGRP :
Le CGRP est surtout localisé dans les parois vasculaires des petites artères musculaires et, en moindre quantité, dans la lame basale et entre les cellules épithéliales.
Son action principale est vasomotrice, avec une action vasodilatatrice de longue durée et un rôle facilitant le remplissage des lacs veineux turbinaux.
Il n’a pas, a priori, d’action sur les secrétions glandulaires.
* Neuropeptide Y :
Libéré par les fibres sympathiques, il a une action vasoconstrictrice mais semble également capable de modifier le volume sécrétoire.
Son application locale chez des sujets sains et allergiques suggère que l’exsudation protéique qui en résulte est due à une modification de la perméabilité vasculaire.
En revanche, il n’a pas d’action sur la sécrétion des glandes sous-muqueuses.
* NEP :
C’est une enzyme régulant l’activité des neuropeptides libérés dans la muqueuse respiratoire.
Son étude, à partir de biopsies de cornet inférieur et de secrétions nasales, montre qu’elle est surtout présente dans les glandes sous-muqueuses, les cellules épithéliales et les cellules myoépithéliales des petits vaisseaux.
Sa présence dans les secrétions nasales est, en revanche, beaucoup plus faible.
D’autres substances semblent capables de stimulation sur les terminaisons nerveuses.
Récemment chez le cochon d’Inde, Sekizawa a obtenu, mais dans des conditions non physiologiques, des réponses sur les terminaisons sensitives du nerf ethmoïdal après stimulation histaminique.
Il en conclut que certaines fibres sensitives seraient stimulables par l’histamine.
L’augmentation de la sensibilité des fibres sensitives à des éléments extérieurs tel que le dioxyde, l’hyperventilation ou l’inhalation d’air froid se traduit par une libération de cytokines (IL1, IL6, IL8, IL11 et TGFalpha) qui entraîne une hyperalgie.
G – APPAREIL MUCOCILIAIRE :
L’épithélium respiratoire est un épithélium pseudostratifié qui comprend des cellules ciliées, des cellules muqueuses et des cellules basales.
Elles reposent sur un tissu de soutien, constituant la lame basale.
La cohésion de l’épithélium est assurée par plusieurs systèmes jonctionnels :
– les jonctions serrées (« zonula occludens ») au niveau apical des cellules ;
– les jonctions intermédiaires (« zonula adherens ») sous les jonctions serrées, assurent la communication entre les cellules adjacentes ;
– les desmosomes (« macula adherens ») reliés entre eux par des filaments de cytokératine, assurent le maintien de la morphologie cellulaire ;
– les jonctions communicantes mettent en communication (2 nm) directe le cytoplasme des cellules adjacentes ; elles participent au transfert de messagers secondaires : calcium, inositol triphosphate ;
– les hémidesmosomes favorisent l’ancrage des cellules à la lame basale ; la lame basale soutient l’épithélium : elle est formée par un réseau complexe de protéines (laminine, protéoglycanes, collagène IV…) ; deux zones sont définies lors de son examen en microscopie électronique : une zone claire, lamina lucida et une zone dense, lamina densa.
Longtemps considérée comme inerte, cette structure a de nombreuses propriétés biologiques.
Elle entretient, par le système des intégrines, d’étroites interactions avec les cellules épithéliales, en particulier dans leur migration, leur prolifération et leur différenciation.
Les cellules de l’épithélium de surface sont ancrées sur la lame basale.
L’ancrage à la lame se fait au moyen de récepteurs membranaires : les intégrines.
Ce sont des récepteurs membranaires aux protéines de la lame basale.
Elles interviennent dans la migration des cellules inflammatoires et facilitent, in vitro, l’adhésion des cellules aux protéines de la matrice extracellulaire.
Elles fonctionnent comme des molécules capables de détecter l’attachement ou le détachement des cellules sur la matrice extracellulaire.
La laminine joue un rôle dans la prolifération cellulaire.
Des glandes s’invaginant à partir de l’épithélium dans la sousmuqueuse sont formées par des cellules séreuses et muqueuses.
Les cellules muqueuses sont caractérisées par la présence de granules sécrétoires clairs aux électrons, renfermant des mucines et des protéines antibactériennes (IgA).
Les cellules séreuses contiennent des granulations intracytoplasmiques denses aux électrons.
Elles synthétisent des glycoprotéines, des protéines à propriété antibactérienne (lactoferrine, lysozyme) et antioxydante (transferrine, antileucoprotéase).
Elles libèrent également des IgA synthétisées par les plasmocytes.
Des cellules myoépithéliales sont également identifiées autour des acini glandulaires.
Elles possèdent les caractéristiques des cellules épithéliales et des cellules musculaires.
Sous la lame basale, dans l’environnement des glandes de la sous-muqueuse, siège la matrice extracellulaire.
Elle est composée de molécules de collagène, d’élastine et de protéoglycanes synthétisées principalement par les fibroblastes.
De nombreux facteurs de croissance sont également présents dans ce réseau de macromolécules, ainsi que quelques cellules inflammatoires et des vaisseaux.
Tout ce réseau cellulaire communique à la fois avec l’environnement extérieur, mais également avec l’ensemble de l’organisme.
Le rôle protecteur de l’épithélium respiratoire est assuré par l’appareil mucociliaire et par les éléments de la sous-muqueuse.
Mucus : première barrière de défense
Le mucus est la première barrière de défense.
Les composants du mucus assurent le piégeage des particules inhalées et leur inactivation par différents mécanismes : antibactériens, antiprotéasique et antioxydant.
Le mucus est composé principalement d’eau (95 %) et d’un réseau macromoléculaire de mucines (4 %).
Deux phases sont décrites.
La première, superficielle, dite « gel », a une viscosité et une élasticité élevées.
La couche profonde, aqueuse, périciliaire, est dite « sol ».
Le mucus est en constant mouvement en raison de l’activité ciliaire des cellules épithéliales, il assure le maintien de l’hydratation de l’épithélium.
Son pH varie de 6,5 à 7,8.
La régulation hydrique est contrôlée par l’absorption d’ions sodium et la sécrétion d’ions chlore.
La sécrétion transépithéliale d’ions et d’eau est régulée par des systèmes actif et passif.
Les principaux systèmes connus sont le cotransporteur Na/K/2CL, l’échangeur Na/K et différents canaux chlorures potassique et sodique.
Les canaux « chlore », situés au pôle apical des cellules respiratoires, ont été particulièrement étudiés en raison de leur rôle dans la mucoviscidose.
Parmi eux, le canal CFTR régulé par la voie de l’AMPc (acide adénosine-monophosphorique cyclique).
La protéine CFTR, codée par le gène situé sur le chromosome 7, constitue elle-même un canal chlore de faible conductance.
Les mucines sont des glycoprotéines de masse moléculaire élevée (1 000 kDA) qui forment un réseau macromoléculaire assurant le piégeage des molécules dont le diamètre est supérieur à 2 mm.
Elles assurent également la neutralisation des micro-organismes par leurs chaînes carbohydrates.
D’autres éléments sont présents dans le mucus.
Le lysozyme, sécrété par les cellules séreuses, possède une activité bactériolytique (Gram+) et stimule l’activité phagocytaire des leucocytes et des macrophages.
Les IgA, synthétisées par les plasmocytes de la sous-muqueuse, sont internalisées dans les glandes séreuses et muqueuses, pour être libérées dans la lumière respiratoire.
Elles inhibent l’adhérence des bactéries, neutralisent les virus dans les cellules et favorisent l’activité phagocytaire des cellules inflammatoires.
Les phospholipides contrôlent la rhéologie du mucus.
La transferrine, glycoprotéine sécrétée par les glandes séreuses, fixe le fer nécessaire à la croissance des bactéries, ce qui assure une protection antibactérienne.
Des inhibiteurs de protéases sont également présents dans le mucus.
Ils préviennent les dommages cellulaires déclenchés lors de pathologies inflammatoires.
Les antioxydants luttent contre les effets des radicaux libres et des molécules oxydantes produits par des agents chimiques, des substances toxiques ou des cellules inflammatoires.
Toutes ces molécules prennent part au maintien de l’homéostasie du mucus dont le transport est assuré par les battements ciliaires des cellules épithéliales à une vitesse de 10 à 15 mm/min.
La fréquence des battements est d’environ 10 à 15 Hz.
Chaque cellule ciliée possède environ 200 cils.
Les cils sont animés de battements périodiques dont les caractéristiques ont été analysées par de nombreux travaux.
Le battement ciliaire est coordonné : tous les cils battent dans la même direction avec la même fréquence de façon synchrone (rythme métachronal).
Chaque cycle de battement comprend deux phases :
– la phase active, deux fois plus brève que la phase de retour, permet au cil de se déployer et d’atteindre sa longueur maximale ; le cil « accroche » le mucus et le propulse ;
– la seconde phase, dite phase de repos, d’environ 10 ms, permet au cil de revenir dans sa position initiale avant la prochaine phase active ; l’énergie nécessaire à ce mouvement est fournie par l’hydrolyse de l’ATP (adénosine triphosphate).
L’analyse ultrastructurale des cils montre que le mouvement ciliaire est la résultante d’un glissement entre les microtubules qui arment le cil : neuf paires périphériques, une paire centrale.
Ce glissement est possible par les modifications des zones d’attache des bras de dynéine sur ces microtubules.
La liaison entre les bras de dynéine et les microtubules est ATP-dépendante.
En présence d’ATP, la liaison est rompue et rétablie en absence d’ATP.
La régulation et la coordination de ces interactions demeurent inconnues.
Les cultures cellulaires épithéliales in vitro ont confirmé que les battements ciliaires persistent plusieurs heures si le milieu nutritif est suffisant.
De nombreux facteurs influencent le transport mucociliaire : température, hygrométrie, tabac, etc.
Une étude avec 185 patients pathologiques et 16 patients témoins suggère une asymétrie dans le transport mucociliaire, mais l’étude n’a pas évalué si cette asymétrie se modifiait avec le temps et en particulier s’il y avait une corrélation dynamique avec l’état de congestion de la muqueuse.
Le monoxyde d’azote (NO) chez l’animal augmente la fréquence du battement ciliaire in vitro.
Chez l’humain volontaire sain, le NO modifie la microcirculation nasale et augmente également l’activité mucociliaire.
Son rôle a été récemment évoqué en pathologie puisque Lundberg a trouvé, chez trois enfants porteurs d’anomalies ciliaires (syndrome de Kartagener), de faibles concentrations de NO dans l’air expiré, ce qui tendrait à confirmer une relation entre la concentration de NO et l’activité cilaire.
La composition cellulaire du mucus est aussi variable ; ainsi, au contact de l’allergène, une augmentation des éosinophiles est signalée.
L’absence de ventilation nasale ne modifie pas le transport mucociliaire.
En revanche, le nombre de cellules glandulaires diminue avec le temps, mais de manière modeste.
Si la substance P augmente le transport mucociliaire chez l’animal, chez l’homme ce phénomène n’est pas trouvé lors d’étude in vitro.
Ces résultats contradictoires, comparés aux études chez l’animal et chez l’humain in vitro, s’expliquent probablement en raison des NEP qui dégradent les NP comme la substance P ; d’ailleurs, une diminution de leur activité semble être à l’origine d’une réponse inflammatoire plus intense et d’une hyperréactivité.
H – SYSTÈME DE DÉFENSE NASOSINUSIEN :
Il est classique de décrire trois systèmes de défense :
– le premier correspond à la barrière épithéliale et au système mucociliaire déjà décrits, associés à la flore commensale des cavités nasales ;
– le second comprend le système immunitaire annexé aux muqueuses respiratoires : l’IgA et le NALT ;
– le troisième correspond aux mécanismes d’inflammation non spécifiques mis en route en cas de rupture des équilibres physiologiques.
1- Système mucociliaire et flore commensale :
L’appareil mucociliaire participe à la défense de l’organisme en éliminant les particules et les micro-organismes captés par le mucus, mais également en participant à la réponse immunitaire par l’intermédiaire des lymphocytes siégeant dans l’épithélium et peut-être par les cellules épithéliales elles-mêmes.
La flore commensale joue également un rôle important dans cette défense.
Elle est variable selon la localisation dans la cavité nasale et varie probablement avec l’âge.
On trouve essentiellement Staphylococcus epidermidis, des corynébactéries, le Staphylococcus aureus est fréquemment signalé. Des anaérobies sont aussi présentes dans le méat moyen.
Ces germes jouent probablement un rôle dans l’équilibre écologique de la cavité nasale et évitent probablement la greffe de micro-organismes plus agressifs (Haemophilus, pneumocoque) qui peuvent toutefois être portés par 3 à 5 % de la population sans pathologie associée.
2- Système immunitaire :
La muqueuse nasosinusienne fait partie de l’ensemble des muqueuses respiratoires. Elle contient des cellules immunitaires appartenant au mucosa-associated lymphoid tissue (MALT) ou tissu lymphoïde associé aux muqueuses.
Ce système est constitué de tissu lymphoïde diffus étroitement lié à l’épithélium et à la sousmuqueuse.
On y trouve tous les éléments cellulaires : cellules présentatrices d’antigènes, lymphocytes T et B, plasmocytes… et des Ig.
* IgA sécrétoire :
L’Ig principale du système est l’IgA dont on évalue la synthèse à 4 g/j dans l’organisme.
La dimérisation de l’IgA est assurée par un peptide synthétisé par les plasmocytes (pièce J).
Le dimère est ensuite capté par la pièce sécrétoire élaborée par les cellules épithéliales.
Par un mécanisme particulier de fusion des membranes, une molécule complexe est formée comprenant deux IgA, une pièce J et une pièce sécrétoire : l’IgA sécrétoire.
L’IgA sécrétoire est produite localement par des cellules immunocompétentes disséminées dans la muqueuse.
Sa production est facilitée par des lymphocytes T.
Chez la souris, la production d’anticorps IgA spécifiques par les cellules immunocompétentes dans le NALT joue un rôle important dans la réponse à l’infection.
Son rôle spécifique est de former de volumineux complexes immuns avec des antigènes spécifiques qui sont ensuite transportés par le mucus vers le tube digestif.
* NALT :
Récemment, le NALT, ou tissu lymphoïde appendu à la muqueuse nasosinusienne, a été individualisé et fait l’objet de nombreux travaux chez l’animal et chez l’homme.
Sa composition et sa localisation sont encore l’objet de controverses.
Avant d’envisager sa description et son fonctionnement, un bref rappel sur les lignées lymphocytaires est proposé.
Les lymphocytes sont différenciés à partir des antigènes de différenciation (cluster of differenciation : CD), des récepteurs reconnaissant l’antigène et des molécules d’adhésion présents sur la membrane.
Les lymphocytes T-helper (CD4+) sont eux-mêmes répartis en deux sous-populations selon leur profil sécrétoire.
Les T-helper 1 synthétisent surtout l’INFç et l’IL2 et réduisent la synthèse d’IgE par les cellules B. Les T-helper 2 synthétisent principalement l’IL4, l’IL5 et l’IL10.
L’IL4 augmente la synthèse d’IgE par les cellules B et l’IL5 amplifie la réaction inflammatoire (éosinophile).
Le rapport CD4+/suppresseur (CD8+) est un élément important dans la réponse muqueuse à l’inflammation.
Sa valeur est différente selon les pathologies envisagées, mais les résultats actuellement publiés sont parfois contradictoires.
Néanmoins, il semble acquis que le nombre de CD4+ est plus important chez le sujet normal.
Une étude récente chez l’animal amène à supposer que le NALT serait constitué, dans la muqueuse nasale, de lymphocytes de phénotype T-helper 0 qui, selon les conditions d’environnement, seraient capables d’acquérir un phénotype de type T-helper 1 et/ou T-helper 2.
Dans la lumière nasale, le phénotype T-helper 2 serait prédominant, favorisant ainsi la production IgA locale.
En cas d’infection, une augmentation des lymphocytes T-helper 1 serait communément admise.
Dans la polypose, une majorité de lymphocytes T avec un grand nombre de CD4+ est identifiée, mais dans une proportion identique à celle du sujet témoin.
Plus récemment, Kamil a étudié la population cellulaire immunocompétente chez le sujet allergique, dans différentes cavités nasosinusiennes.
La densité en éosinophiles était plus importante dans l’ethmoïde par rapport au sinus maxillaire luimême, plus riche que le cornet inférieur.
La valeur du rapport CD4+/CD8+ était également plus importante dans l’ethmoïde que dans le sinus maxillaire et dans le cornet inférieur.
Pour les cytokines, l’IL4 était plus fréquente dans le cornet inférieur où siègent en majorité des mastocytes.
À l’inverse, l’IL5 a été trouvée en grande quantité dans l’ethmoïde et le sinus maxillaire, probablement sécrétée par les CD4+.
L’auteur souligne d’ailleurs le rôle prépondérant du rapport CD4+/CD8+ pour expliquer la répartition hétérogène des éosinophiles.
Il s’interroge également sur le risque qu’entraînerait chez l’allergique certaines techniques chirurgicales visant à élargir la communication du sinus maxillaire avec les cavités nasales, en augmentant l’exposition de la muqueuse maxillaire à l’environnement et donc à une hyperstimulation antigénique.
Dans l’allergie nasale, le système immunitaire nasal semble jouer un rôle de plus en plus prépondérant.
L’allergène, le plus souvent un pneumallergène, est capté par le mucus et franchit la muqueuse.
Il est pris en charge par les macrophages et les cellules accessoires qui présentent sur leur surface des molécules de classe II du système majeur d’histocompatibilité (HLA-DR+).
D’autres cellules présentent ce marqueur : cellules épithéliales, lymphocytes B et T, fibroblastes, mais leur rôle est encore méconnu.
Une fois traité par les cellules présentatrices, l’antigène est présenté aux lymphocytes ; un complexe se forme avec les lymphocytes T à l’aide des molécules d’adhésion qui stabilisent la liaison CD3- récepteur T-molécule HLA.
Le complexe CD3-récepteur T (ou T-cell receptor [TCR]) est composé de deux chaînes : soit alpha-bêta (90 %), soit gamma-delta (10 %).
Une fois l’allergène fixé sur le récepteur lymphocytaire T, une activité enzymatique est déclenchée aboutissant à la mobilisation du calcium intracellulaire et à une activation de gènes qui permettront une différenciation fonctionnelle du lymphocyte activé.
Par ailleurs, on observe une augmentation du nombre de T-helper 2 qui vont libérer l’IL4, facteur de croissance pour les lymphocytes et induire la transformation des T-helper 0 en T-helper 2.
D’autres cellules, comme les basophiles et les mastocytes, produisent également de l’IL4.
Les T-helper 2 favoriseraient la synthèse d’IgE par l’intermédiaire d’un contact direct entre les lymphocytes B et T.
Les lymphocytes B peuvent, en outre, être stimulés directement par leurs immunoglobulines de surface par l’antigène sous sa forme native.
Une fois activée, la formation des lymphocytes B spécifiques d’un antigène se ferait dans le ganglion et peut-être directement dans la muqueuse nasale.
Outre ses actions spécifiques précédemment décrites, les cellules du NALT semblent jouer un rôle dans la régulation du système IgA.
Une meilleure connaissance de ce système laisse également entrevoir des possibilités de vaccination par stimulation intranasale, déjà à l’étude, en particulier chez l’animal, avec des résultats encourageants.
* Réaction inflammatoire non spécifique :
Elle est déclenchée par la rupture des équilibres physiologiques.
Elle peut être de mécanisme immun ou non.
Plusieurs systèmes de défense peuvent être mis en jeu isolément ou en même temps.
À la suite d’une lésion tissulaire, quelle qu’en soit la nature, la première étape de l’inflammation est l’activation du facteur XII par la voie classique et du complément par la voie classique ou alterne.
Lors d’une blessure endothéliale, les plaquettes sont activées et provoquent le relargage des facteurs de la coagulation et de l’inflammation.
Enfin, un nombre important de stimuli, spécifiques ou non, activent le mastocyte, véritable cellule sentinelle.
La libération histaminique qui en résulte est à l’origine d’une cascade de réactions qui vont amplifier la réaction inflammatoire par son action directe sur les vaisseaux et sur les fibres nerveuses.
Lors des réactions immunes, le fragment C1q (par la voie active) reconnaît le fragment Fc de l’anticorps (IgM, IgG1, IgG2, IgG3), entraînant une cascade de réactions pour aboutir à une action cytotoxique.
Toutes ces réactions ont un objectif : combattre l’agression quelle qu’elle soit.
Les modifications de la microcirculation ont un rôle fondamental, en favorisant l’arrivée des cellules inflammatoires sur le site qui vont infiltrer le tissu interstitiel.
La nature et le nombre de ces cellules varient selon l’état immun du patient et le type d’agression. Les médiateurs libérés sont également, en quantité et en nature, sous la dépendance des mêmes paramètres.
Les facteurs régulant la venue de ces cellules et leur profil sécrétoire restent hypothétiques.
Toutefois, récemment, des travaux ont permis de proposer un mécanisme à l’origine de la migration des cellules éosinophiles dans la muqueuse nasale lors de stimulation antigénique.
Denburg a largement contribué à promouvoir cette hypothèse.
Il a montré, d’abord chez l’animal, qu’après une stimulation antigénique, une augmentation des cellules progénitrices des éosinophiles survenait dans la circulation sanguine.
Une seconde expérience lui a permis de confirmer ce résultat, mais également d’obtenir une augmentation de ces cellules directement dans la moelle osseuse.
À partir de ces travaux préliminaires, il a obtenu des résultats similaires pour des sujets asthmatiques après stimulation antigénique.
Ces travaux encore en cours suggèrent une interaction importante entre la muqueuse nasale et la moelle osseuse par l’intermédiaire de médiateurs encore non précisément étudiés.
Les résultats de ces études expliqueraient également, en partie, la venue massive des cellules inflammatoires lors d’une aggression muqueuse.
En revanche, les signaux responsables de la domiciliation nasale de ces cellules restent totalement inconnus.
+ Phase de restauration :
Une fois tous les acteurs en place, un mécanisme de régulation et d’inactivation va permettre le nettoyage et la restauration des tissus lésés.
Ces mécanismes sont complexes et encore peu étudiés, on y trouve de nombreuses actions enzymatiques (antileucoprotéases, kininases, endopeptidases, etc) et cellulaires (fibroblastes, macrophages, lymphocytes, etc).
Symptomatologie fonctionnelle nasale :
Si les fonctions nasales peuvent être maintenues ou restaurées, dans des conditions extrêmes parfois, soit transitoirement, soit définitivement apparaissent des dysfonctions nasales.
L’obstruction nasale, les troubles de l’odorat, la rhinorrhée, les éternuements, les algies faciales sont les principaux symptômes fonctionnels signalés par les patients consultant pour un problème nasal.
A – OBSTRUCTION NASALE :
Elle correspond à une sensation subjective de gêne à l’écoulement de l’air dans les cavités nasales.
Aucun examen objectif ne permet de quantifier cette gêne nasale.
Le clinicien est obligé de se contenter des résultats de l’interrogatoire pour évaluer la sensation nasale.
Toutefois, deux examens peuvent l’aider à quantifier le débit de l’air qui traverse la cavité nasale (rhinomanométrie) et les zones de rétrécissement que rencontre l’air lors de son passage dans la cavité nasale (rhinométrie acoustique).
1- Rhinomanométrie :
Elle permet la mesure simultanée du débit et des variations de pression du courant aérien traversant les cavités nasales.
Les règles de sa pratique sont proposées par le comité international de standardisation de la rhinomanométrie.
La rhinomanométrie antérieure active est l’examen le plus accessible au consultant.
La mesure de la résistance nasale correspond au rapport de la différence de pression entre l’entrée et la sortie de la cavité nasale sur le débit en cas de flux laminaire (soit inférieur ou égal à 150 mL/s).
Elle s’exprime en Pa/mL/s, soit 0,30 Pa/mL/s.
L’examen est réalisé chez le sujet au repos sans préparation, avec vasoconstriction, en respiration calme.
De nombreuses autres manoeuvres peuvent être effectuées (effort, décubitus, etc) ; le résultat est habituellement représenté sous la forme d’un graphique.
Cette mesure de la résistance nasale a un intérêt diagnostique (allergie professionnelle), mais surtout comparatif chez un même patient pour vérifier l’efficacité d’une thérapeutique médicale ou chirurgicale.
2- Rhinométrie acoustique :
Elle étudie la réflexion d’une impulsion acoustique à l’intérieur des cavités nasales.
Le son recueilli est traité informatiquement pour obtenir le calibre de la cavité nasale sur toute la propagation de l’onde sonore.
La pratique de cet examen se heurte à sa sensibilité et au manque de standardisation de cette technique.
La courbe obtenue après plusieurs stimulations sonores reproduit les obstacles rencontrés par le son.
Son emploi reste limité, en France, à quelques équipes, probablement en raison du coût de l’appareil et des trop nombreuses discordances signalées dans la littérature.
Ainsi, l’interrogatoire et l’examen endonasal, couplés à ces examens, peuvent guider le clinicien dans la recherche de l’obstacle à la ventilation nasale qui peut être morphologique (déviation septale), fonctionnel (inflammation) ou mixte.
B – RHINORRHÉE :
Si la production de mucus en condition normale ne se traduit par aucun symptôme ni nécessité d’un mouchage, une production excessive et/ou un trouble de son transport ou de sa réabsorption se traduisent par une rhinorrhée.
Les modifications du mucus peuvent toucher son volume, sa viscosité, son contenu cellulaire.
L’analyse des rhinorrhées souffre d’une absence de consensus dans son mode de recueil et son analyse.
Seuls peut-être la présence d’une éosinophilie est signalée par la plupart des auteurs comme un élément diagnostique dans la recherche d’une rhinite inflammatoire allergique ou non.
La normalité du transport mucociliaire peut néanmoins être appréciée en pratique par des tests simples, soit à l’aide de colorant ou de saccharine.
Ces tests ne permettent toutefois pas d’identifier la cause de l’anomalie mucocilaire.
C – ÉTERNUEMENTS :
Les terminaisons nerveuses du nerf trijumeau sont les récepteurs de l’arc réflexe dont le mécanisme n’est pas totalement connu.
La zone de déclenchement semble siéger dans la région des méats moyen et supérieur.
L’effet protecteur des antihistaminiques suggère le rôle important de l’histamine dans cette réaction, toutefois bien d’autres stimulations peuvent intervenir : physique, chimique, etc.
D – DOULEURS FACIALES :
Comme l’obstruction nasale, elles demeurent un symptôme où la subjectivité est importante.
Si la douleur de la sinusite est unanimement acceptée, les douleurs d’origine nasale restent un sujet de polémique tant dans leur existence que dans leur physiopathologie.
Les travaux, peu nombreux d’ailleurs sur ce sujet, restent contradictoires et les résultats de traitements proposés peu convaincants.
Il convient dans tous les cas de rester très prudent dans la démarche diagnostique et de s’assurer d’un suivi assez long pour confirmer un résultat immédiat, souvent encourageant mais peu stable avec le temps.
Conclusion :
L’organe nasal est un organe encore mystérieux, bien qu’à la portée des cliniciens et des chercheurs.
Ses diverses fonctions, régulées par des mécanismes précis et fiables, sont conservées ou rétablies dans des conditions parfois extrêmes.
L’augmentation sensible des troubles rhinosinusiens justifie que le clinicien s’intéresse de plus en plus à la physiologie nasale pour en connaître les possibles dérèglements et y apporter une thérapeutique précise et adaptée.







")

")
")
{kind=link}