1. Puisque les mutants ne peuvent pas pousser en milieu complet à 42 °C, alors qu’ils
peuvent pousser sur milieu minimum à 36 °C, cela prouve qu’ils ne sont vraisemblablement
pas touchés dans un gène du métabolisme (en général compensable en milieu complet) mais
plutôt dans un gène, noté m, impliqué dans une fonction vitale du cycle cellulaire.
2. La première hypothèse, H1, est toujours possible; il s’agit en général d’une mutation fauxsens
conduisant, par la substitution d’un acide aminé par un autre, à un produit actif mais
thermoinstable.
La deuxième hypothèse, H2, n’est possible qu’en raison du protocole utilisé : le mutant
obtenu a perdu une fonction vitale par suite de l’apparition d’un codon stop (ambre) dans la
séquence codante d’un gène m vital, et ne peut être sélectionné que si l’effet de cette mutation
est supprimé par le suppresseur informationnel d’ambre déjà présent avant la mutagenèse.
Pour mettre en évidence l’existence de la mutation stop (à travers son effet létal) il faut que
le suppresseur soit conditionnel, ce qui est le cas ici puisqu’il est thermosensible.
Le but de ce protocole est, précisément, de pouvoir cribler des mutants non-sens de gènes
impliqués dans des fonctions vitales du cycle cellulaire.
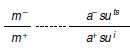
3. a Sous l’hypothèse H1 le mutant m1 a pour génotype (mts ; a–, suts) où mts est la mutation
thermosensible du gène touché lors de la mutagenèse. Dans ce cas, le diploïde obtenu dans le
croisement avec sauvage s’écrira :
Toutes les spores parentales ou recombinées sont ici capables de pousser sur milieu complet
à 36 °C, ce qui n’est pas compatible avec l’observation de tétrades contenant une ou deux
spores qui en sont incapables.
Le mutant m1 n’est pas porteur d’une mutation thermoinstable
(hypothèse 1).
Sous l’hypothèse H2, le mutant m1 a pour génotype (m–; a–, suts) où m– est une mutation
non-sens du gène m touché lors de la mutagenèse, mais dont l’effet peut être supprimé par le
suppresseur suts. Dans ce cas, le diploïde obtenu dans le croisement avec sauvage s’écrira :
Les spores recombinées de génotype (m–; a+, sui) sont incapables de pousser sur milieu
complet à 36 °C en raison de la séparation, par recombinaison, entre m– et le suppresseur
actif.
Le mutant m1 est donc un mutant non-sens… par mutation ambre puisque son effet est
corrigé par le suppresseur d’ambre de a–.
3. b On a 19 ditype-parentaux, 17 ditypes recombinés et 4 tétratypes, ce qui conduit à la
conclusion que les locus de m et su sont génétiquement indépendants (f [DP] = f [DR]) et
physiquement indépendants (f [T] < 2/3).
4. Il suffit de fragmenter par digestion partielle le génome de B et d’insérer les fragments en
un site adéquat du plasmide (en général une séquence polylinker clivée par le même enzyme
que celui qui a servi à la digestion partielle de B).
On amplifie les plasmides recombinants par infection d’une souche de coli étalée sur Mo
+ ampicilline.
On extrait les plasmides et on transfecte le mutant m1 qu’on étale sur milieu
Mo à 42 °C.
Le suts ne peut fonctionner et les gènes m et a ne sont pas exprimables, sauf si le
plasmide apporte le suppresseur d’ambre thermostable de la souche B.
Exercice 8.6
Le gène GAL4 de Saccharomyces cerevisiae code pour un activateur de
certains des gènes de métabolisation du galactose GAL7, GAL10, GAL1,
GAL2; ces quatre gènes possèdent, dans leur promoteur, une séquence
UAS (Upstream Activating Sequence) cible de la protéine PGAL4 qui n’est
activatrice qu’en présence de galactose, celui-ci jouant donc le rôle
d’inducteur.
La protéine PGAL4 possède deux domaines physiquement distincts, aux
fonctions elles-mêmes distinctes, le domaine DBD (DNA Binding Domain),
dans la partie Nter, sert à la liaison de GAL4 à l’ADN sur la séquence UAS;
l’autre domaine AD (Activating Domain), dans la partie C-ter, interagit avec
le complexe de transcription pour activer celui-ci.
Par biologie moléculaire, on a pu dissocier la séquence du gène GAL4 et
cloner dans des vecteurs séparés la séquence DBD et la séquence AD.
L’utilisation de ces deux séquences est à l’origine du premier système de
criblage de gène par la technique des « doubles hybrides ».
De nombreux gènes sont impliqués dans le cycle cellulaire, ils codent pour
des produits qui interagissent entre eux et forment des complexes multimériques
permettant le blocage du cycle dans une phase, ou sa transition vers
la phase suivante.
Parmi ces gènes, le gène CDK2 (Cycline-Dependent
Kinase) a été cloné et code pour une kinase impliquée spécifiquement dans
la transition G1/S, en association avec le produit d’un autre gène encore
inconnu.
On souhaite cloner et identifier le (ou un) gène codant pour la (ou une)
protéine partenaire de CDK2 et on dispose pour cela de :
– une souche A de levure, délétée pour les gènes URA3 et LYS2, et porteuse
de la séquence codante du gène LACZ de coli sous la dépendance d’un
promoteur contenant une séquence UAS spécifique de PGAL4.
L’expression du gène LACZ permet la synthèse de β-galactosidase dont
la présence peut être phénotypiquement attestée par la coloration bleue
des colonies quand le milieu contient du X-GAL;
– un plasmide B porteur du gène de sélection sauvage URA3, et de la
séquence DBD de GAL4 clonée en phase avec la séquence du gène CDK2
(le domaine DBD étant en extrémité N-ter de la protéine de fusion);
– un plasmide C porteur du gène de sélection sauvage LYS2 et de la
séquence AD de GAL4, en amont de laquelle existe un site d’insertion qui peut, par exemple, permettre de cloner en phase la séquence d’un ADNc (le domaine AD étant en extrémité C-ter de la protéine de fusion);
– des boîtes de milieu minimum additionné de X-GAL.
La première étape du protocole consiste à extraire des ARNm de cellules
d’une culture en croissance exponentielle puis de les transformer en ADNc
pour cloner ceux-ci dans des plasmides C.
Après avoir justifié cette première étape, vous proposerez la suite du protocole.
Sélection d’un gène par double-hybride.
Solution
• On cherche à isoler un gène spécifiant une protéine partenaire du produit de CDK2; elle est
certainement présente dans des cellules en division puisqu’elle est impliquée dans le passage
en phase S et on doit donc obtenir, par extraction et purification, des messagers de ce gène à
partir desquels des ADNc pourront être synthétisés puis clonés en phase avec la séquence AD,
dans les plasmides C.
• Des colonies de la souche A ne peuvent se développer sur des boîtes de milieu minimum si
les cellules n’ont pas été, au préalable, cotransformées par le plasmide B qui apporte URA3
et le plasmide C qui apporte LYS2.
Le plasmide B permet la présence d’un produit de fusion DBD-CDK2 qui se fixera sur les
séquences UAS-GAL4, dont celle du gène LACZ, mais il n’y aura pas de transcription de
celui-ci en absence de la séquence AD.
La séquence AD est présente dans la cellule, dans une protéine de fusion P-AD, où P est la
protéine spécifiée par l’ADNc cloné dans le plasmide C acquis par la cellule. Mais, ne disposant
pas de domaine DBD, cette protéine de fusion P-AD ne peut stimuler l’expression
d’aucun gène.
À moins que la protéine P soit précisément le partenaire de CDK2, on peut alors supposer
(espérer, et cela marche) que l’association des deux protéines de fusion DBD-CDK2/P-AD
va constituer un dimère porteur du domaine DBD en N-ter et AD en C-ter, capable de
stimuler la transcription de LACZ aussi bien, ou presque, que le ferait la protéine GAL4.
Alors le gène reporter LACZ sera exprimé, et la colonie sera bleue.
• On transforme donc des cellules de la souche A avec les plasmides B et C, ceux-ci ayant
cloné un ADNc, et on étale sur des boîtes de milieu Mo + X-GAL.
On sélectionne la ou les
colonies bleues qui sont susceptibles de contenir l’ADNc d’un gène spécifiant un partenaire
du produit codé par CDK2.
On remarquera que ce gène, cloné par la technique des doubles-hybrides (deux protéines de
fusion ou hybrides s’assemblant pour former l’activateur d’un gène reporter), aura été identifié
et cloné sans qu’on soit passé par sa mutation et les étapes de la génétique classique.