En croisant deux souches pures de pois à fleurs pourpres et à fleurs blanches,
on a obtenu des descendants F1 homogènes à fleurs pourpres.
Le
croisement F1 × F1 a fourni 320 plantes F2, 142 à fleurs blanches et 178 à
fleurs pourpres.
Quelle est l’interprétation la plus simple de ces résultats ?
– Modification des proportions classiques de ségrégation dans le cas d’intéraction
entre gènes.
– Exemple de
la couleur de la fleur chez le pois.
Solution. Le phénotype blanc est récessif.
Il n’y a pas ségrégation 2 × 2 chez la F1; en effet
les proportions observées divergent significativement des proportions attendues pour un
couple d’allèles, soit 3/4-1/4, ce qui donnerait, pour un effectif total de 320 plantes, 240 à
fleurs pourpres et 80 à fleurs blanches.
Un test de χ2 donne une valeur de [(178 – 240)2/240] + [(142 – 80)2/80] = 48,05, hautement
significative puisque très supérieure à la valeur seuil, au risque de 5 %, d’un χ2 à 1 degré de
liberté, égale à 3,84.
Les souches étudiées diffèrent donc pour plus d’un gène.
Comme le croisement F1 × F1 ne laisse apparaître en F2 que les deux phénotypes parentaux
et que le phénotype à fleur blanche est récessif, on peut supposer que la souche pure de
phénotype récessif est homozygote mutée pour, au moins, deux gènes.
Dans le cas de l’hypothèse minimaliste de deux gènes mutés, la souche blanche est de génotype
(a//a; b//b) alors que la souche pourpre est homozygote {A//A; B//B}.
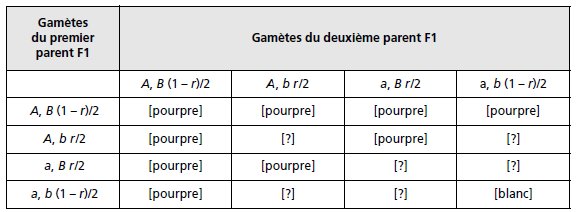
La F1 est un double hétérozygote {A//a; B//b} dont la méiose fournit quatre types de gamètes
dont les fréquences dépendent du taux de recombinaison entre les deux gènes (tabl. 3.21).
Compte tenu des souches de départ et des observations de dominance en F1, il est possible
d’attribuer un phénotype à dix des seize génotypes possibles, puisque la présence simultanée
d’un allèle A et d’un allèle B conduit à la couleur pourpre, et que leur absence simultanée
conduit à la couleur blanche.
TABLEAU 3.21 GÉNOTYPES ET PHÉNOTYPES DE LA F2
APRÈS UNION DES GAMÈTES ISSUS DE LA MÉIOSE CHEZ LA F1.
Plusieurs hypothèses peuvent être formulées pour les autres phénotypes et doivent être validées
ou rejetées en fonction de leur cohérence avec les résultats observés.
– Si les plantes à fleurs blanches sont strictement de génotype double mutant {a//a; b//b} et
que les deux gènes sont indépendants, on s’attend à n’en observer que 1/16 (0,0625), ce
qui n’est pas le cas (142/320 = 0,443 ~ 4/7).
– Si, pour avoir le phénotype à fleurs blanches, il suffit d’être homozygote a//a ou b//b, et
que les gènes sont indépendants, on s’attend à avoir 9/16 [pourpre] et 7/16 [blanche], ce
qui est le cas ici (voir exercice 3.3).
Dans cette hypothèse, les deux gènes ont des effets phénotypiques « de même nature », ils
participent tous deux à la coloration de la fleur et la perte de fonction de l’un des gènes est
phénotypiquement équivalente à la perte de fonction de l’autre.
Cependant, ils n’ont pas la
même fonction, sinon il suffirait d’avoir A ou B pour être coloré; ils ont donc un « effet
complémentaire » dont la somme permet la coloration (c’est le cas des gènes qui agissent
dans la succession des étapes d’une chaîne de biosynthèse).
Mais il faut envisager toutes les hypothèses avec deux gènes afin de vérifier si cette hypothèse,
bien que conforme aux résultats, est la seule possible.
– Si les plantes à fleurs blanches sont strictement de génotype double mutant {a//a; b//b} et
que les deux gènes sont liés, on s’attend à en observer plus que 1/16 (0,0625), ce qui est le
cas (142/320 = 0,443 ~ 4/7).
Dans ce cas, tous les autres génotypes sont de couleur pourpre et il suffirait, sur le plan
fonctionnel de n’avoir que A ou B pour être pourpre; les deux gènes auraient alors la même
fonction (ce qui est le cas quand on a des gènes redondants). Les fréquences de ces deux
phénotypes s’écrivent :
f [pourpre] = (3 – 2r + r2)/4 + r(2 – r)/2;
f [blanc] = (1 – r)2 /4
d’où on tire r = – 0,333, ce qui est incohérent sur le plan théorique, un taux de recombinaison
ne pouvant être négatif.
Une autre démonstration consiste à dire que r étant compris entre 0 et 1/2, la fréquence des
plantes à fleurs blanches ne peut être comprise, sous cette hypothèse, qu’entre 1/4 (0,25) et
1/16 (0,0625), et que la valeur observée de 0,44 est hors de cet intervalle. Cette hypothèse
conduit à une incohérence théorique et doit être rejetée, ce qui valide l’hypothèse
précédente.
Conclusion : Quand deux gènes indépendants ont des effets complémentaires sur un même
phénotype, on observe chez la F2 issue du croisement F1 × F1, une modification des proportions
9-3-3-1 en un regroupement 9-7.
Exercice 3.6
La bourse à pasteur est une plante qui tire son nom de ses fruits en forme
de bourse oblongue.
On connaît deux variétés constituant des souches
pures; la première présente un fruit rond et renflé (bourse pleine) alors que
la seconde présente un fruit plus aplati (bourse vide).
En croisant ces deux souches pures entre elles, on obtient des descendants F1
homogènes [bourse pleine].
Le croisement F1 × F1 a fourni 320 plantes F2,
18 à [bourse vide] et 302 [bourse pleine].
Quelle est l’interprétation la plus
simple de ces résultats ?
➤ Niveau Licence/Définition des objectifs.
– Modification des proportions classiques de ségrégation dans le cas d’intéraction
entre gènes.
– Exemple de
la bourse à pasteur.
Solution. Le phénotype [bourse vide] est récessif.
Il n’y a pas ségrégation 2 × 2 chez la F1, en effet les proportions observées divergent significativement
des proportions attendues pour un couple d’allèles, soit 3/4-1/4; ce qui donnerait,
pour un effectif total de 320 plantes, 240 [bourse pleine] et 80 [bourse vide].
Un test de χ2 donne une valeur de [(302 – 240)2/240] + [(18 – 80)2/80] = 64,06 hautement
significative puisque très supérieure à la valeur seuil, au risque de 5 %, d’un χ2 à 1 degré de
liberté, égale à 3,84. Les souches étudiées diffèrent donc pour plus d’un gène.
Comme le croisement F1 × F1 ne laisse apparaître en F2 que les deux phénotypes parentaux
et que le phénotype à fleur blanche est récessif, on peut supposer que la souche pure de
phénotype récessif est homozygote mutée pour, au moins, deux gènes.
Dans le cas de l’hypothèse minimaliste de deux gènes mutés, elle est de génotype (a//a; b//b)
alors que la souche [bourse pleine] est homozygote {A//A; B//B}.
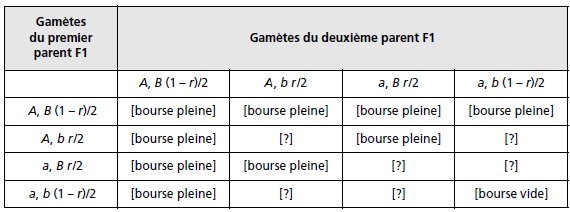
La F1 est un double hétérozygote {A//a; B//b} dont la méiose fournit quatre types de gamètes
dont les fréquences dépendent du taux de recombinaison entre les deux gènes (tabl. 3.22).
Compte tenu des souches de départ et des observations de dominance en F1, il est possible
d’attribuer un phénotype à dix des génotypes possibles, puisque la présence d’un allèle A et
d’un allèle B, chez la F1, conduit au phénotype [bourse pleine] et que leur absence simultanée
conduit au phénotype [bourse vide].
TABLEAU 3.22 GÉNOTYPES ET PHÉNOTYPES DE LA F2
APRÈS UNION DES GAMÈTES ISSUS DE LA MÉIOSE CHEZ LA F1.
Plusieurs hypothèses peuvent être formulées pour les autres phénotypes et doivent être validées
ou rejetées en fonction de leur cohérence avec les résultats.
– Si les plantes de phénotype [bourse vide] sont strictement de génotype double mutant
{a//a ; b//b} et que les deux gènes sont indépendants, on s’attend à n’en observer que
1/16 (0,0625), ce qui est peut être le cas (18/320 = 0,056).
Dans ce cas, tous les autres génotypes sont de phénotype [bourse pleine] et il suffirait, sur
le plan fonctionnel de n’avoir que A ou B pour être de phénotype [bourse pleine]; les deux
gènes auraient alors la même fonction, ce qui est le cas quand on a des gènes redondants ou
dupliqués.
On peut tester, en calculant un échantillon théorique de 320 plantes, la conformité des
fréquences observées par rapport aux valeurs théoriques 15/16 et 1/16.
Le test de χ2 donne une valeur égale à [(302 – 300)2/300] + [(18 – 20)2/20] = 0,21, non
significative puisque très inférieure à la valeur seuil, au risque de 5 %, pour un χ2 à 1 degré
de liberté, soit 3,84.
Rejeter l’hypothèse de conformité serait prendre un grand risque d’erreur; on accepte donc
cette hypothèse génétique mais il convient, cependant, de vérifier qu’une autre hypothèse
n’est pas possible.
– Si, pour avoir le phénotype [bourse vide], il suffit d’être homozygote a//a ou b//b, et que
les gènes sont indépendants, on s’attend à avoir 9/16 [pourpre] et 7/16 [blanche], ce qui
n’est pas le cas.
Conclusion : Quand deux gènes dupliqués ou redondants, mais indépendants, ont des effets
identiques sur un même phénotype, on observe dans le croisement F1 × F1, une modification
des proportions 9-3-3-1 en un regroupement 15-1.
Exercice 3.7 :
On distingue, chez la digitale pourpre, de nombreuses variétés présentant
un phénotype pourpre, blanc, ou magenta; une couleur nettement plus
claire que le pourpre.
On dispose de deux souches pures, A de phénotype [pourpre] et B de
phénotype [blanc]. Le croisement A × B donne des F1 de phénotype
[blanc].
Le croisement F1 × F1 donne des descendants F2, dans les proportions
suivantes, que vous interpréterez : phénotype [blanc] : 12/16; phénotype
[pourpre] : 3/16; phénotype [magenta] : 1/16.
➤ Niveau Licence (L3)/Définition des objectifs.
– Modification des proportions classiques de ségrégation dans le cas d’intéraction
entre gènes indépendants.
– Exemple de la digitale pourpre (Digitalis purpurea).
Solution. Le phénotype [blanc] est dominant.
L’apparition en F2 d’un phénotype non
parental permet de conclure que les deux souches diffèrent par plus d’un gène.
Les proportions phénotypiques 12-3-1 laissent à penser qu’il peut s’agir de deux gènes
indépendants conduisant, dans le croisement F1 × F1, à un rapport 9-3-3-1 modifié en
raison d’un regroupement phénotypique de deux classes correspondant l’une au 9/16, et
l’autre au 3/16 classiquement observés lorsque deux gènes gouvernent des phénotypes
différents.
Ici les deux gènes gouvernent la coloration des fleurs de digitale et leur action n’est ni
complémentaire (on aurait un rapport 9-7), ni redondante (on aurait 15-1).
Les souches étant pures et homozygotes pour des allèles différents, on peut écrire ainsi leurs
génotypes :
– souche A : {a1//a1; b1//b1};
– souche B : {a2//a2; b2//b2}.
Remarque. Bien que le phénotype [blanc] semble dominant, il est imprudent, du fait
de l’interaction entre les gènes, de considérer que la souche B est homozygote pour
un allèle dominant dans chacun de ses deux gènes.
On utilise volontairement une
notation a1 ou a2 qui indique l’origine de l’allèle sans préjuger de l’effet dominant ou
récessif de celui-ci vis-à-vis de l’allèle de l’autre souche.
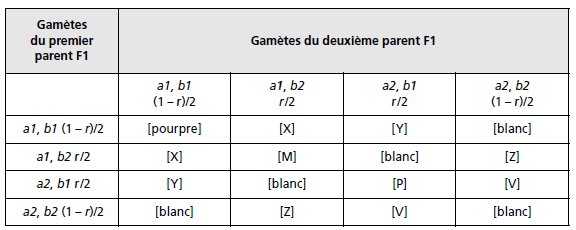
Les descendants F1 seront de génotype {a1//a2; b1//b2}, les F2 issus du croisement F1 × F1,
présenteront divers génotypes détaillés par le tableau 3.23 de croisement des gamètes.
Connaissant le phénotype des souches parentales et de la F1, il est facile de compléter le
tableau pour les génotypes correspondants.
Il reste à définir les phénotypes [M], [P], [X],
[Y], [Z] et [V] correspondant aux nouveaux génotypes générés par la recombinaison
génétique.
Puisqu’on observe seulement 1/16 de [magenta], ce phénotype correspond à [M] ou à [P].
Dans le premier cas, connaissant le génotype de la souche A, on doit conclure que le génotype
a1//a1 conduit au phénotype [pourpre] si la fleur est b1//b1 pour l’autre gène, et au
phénotype [magenta] si elle est b2//b2.
Dans le deuxième cas, connaissant le génotype de la
TABLEAU 3.23 GÉNOTYPES ET PHÉNOTYPES DE LA F2
APRÈS UNION DES GAMÈTES ISSUS DE LA MÉIOSE CHEZ LA F1.
souche A, on doit conclure que le génotype b1//b1 conduit au phénotype [pourpre] si la fleur
est a1//a1 pour l’autre gène, et au phénotype [magenta] si elle est a2//a2.
En fait ces deux cas sont parfaitement symétriques, il suffit de remplacer formellement a
par b et réciproquement.
Considérons donc que nous sommes dans la première situation
[M] = [magenta]; il convient maintenant d’obtenir une interprétation cohérente pour le reste
des résultats, autant sur le plan quantitatif (valeurs des proportions observées) que sur le plan
fonctionnel (effet d’interaction entre les gènes).
Le phénotype [P] du génotype {a2//a2; b1//b1} ne peut pas être [pourpre], puisqu’avec le
phénotype parental, leur somme ne représenterait que 2/16 et que, sommé avec l’un quelconque
des quatre autres phénotypes [X], [Y], [Z] ou [V], la somme des phénotypes
[pourpre] représenterait 4/16, alors que la fréquence observée n’est que de 3/16.
Le phénotype
[P] du génotype {a2//a2; b1//b1} ne peut qu’être [blanc].
Il faut donc déterminer lequel des quatre phénotypes [X], [Y], [Z] ou [V] est le phénotype
[pourpre], en fonction de l’interaction fonctionnelle entre les deux gènes (tabl. 3.24).
TABLEAU 3.24 INTERACTION FONCTIONNELLE ENTRE LES GÈNES ET CORRESPONDANCE GÉNOTYPE/PHÉNOTYPE.
Le parent et le F2 [blanc] sont tous les deux a2//a2 alors que l’un est b2//b2 et l’autre b1//b1;
le gène a semble donc, par son génotype a2//a2, responsable de la couleur blanche (ou
absence de coloration).
Dans ce cas les deux allèles b1 et b2 du gène b gouvernent respectivement la couleur pourpre
et la couleur magenta; le parent b1//b1 et le F2 b2//b2 sont respectivement de couleur
pourpre et de couleur magenta parce qu’au niveau du gène a, ils sont tous deux a1//a1 et non
a2//a2.
Les différences génétiques au niveau du gène b ne s’expriment phénotypiquement que si le
gène a présente un certain génotype, ici a1//a1.
On dit que le gène a exerce un effet épistatique sur le gène b, et plus précisément que l’effet épistatique du gène a est exercé par son
allèle a2.
La question est alors de savoir si l’effet épistatique de l’allèle a2 est récessif ou dominant.
Le
fait que la F1 soit de phénotype [blanc] montre que l’effet épistatique de a2 est dominant.
Si on considère que l’allèle a2 a un effet dominant, tous les génotypes a2//a2 comme a1//a2
seront blancs. Dans ce cas les phénotypes [Y], [Z] et [V] seront blancs et seul le phénotype [X]
sera coloré; ce qui conduira effectivement à 12/16 de [blanc].
Comme les phénotypes [X] sont colorés et de type [pourpre], puisqu’on observe seulement
1/16 de phénotype [magenta] représenté par le phénotype [M], on doit en déduire que l’allèle b1
a un effet dominant sur l’allèle b2, ce qui ne pouvait être mis en évidence dans la F1 du fait
qu’elle était a1//a2 et que l’effet épistatique dominant de a2 bloquait la coloration, et conduisait
à un phénotype [blanc].
On peut donc réécrire les génotypes des deux souches, en utilisant des notations appropriées
aux effets de dominance des allèles :
– souche A : {a//a; B//B};
– souche B : {A//A; b//b};
où le couple d’allèles B-b gouverne respectivement les couleurs pourpre-dominant et
magenta-récessif, et le couple A-a gouverne l’absence de coloration-dominante et la présence
de coloration-récessive.
Remarque. Si on avait choisi le phénotype [P] pour magenta, on serait arrivé aux
mêmes conclusions, avec l’inversion des gènes a et b; le premier gouvernant alors la
couleur et le second la coloration.
Conclusion : Quand deux gènes sont indépendants et que l’un exerce un effet épistatique
dominant sur l’autre, on observe dans le croisement F1 × F1 une modification des proportions
9-3-3-1 en un regroupement 12-3-1.