Recombinaison génétique, indépendance ou liaison génétique, cartographie des gènes
A - Considérations générales
:
Lorsque l’étude de la méiose chez un hétérozygote F1 issu du croisement de deux
souches pures conduit à l’absence de ségrégation 2/2, on conclut évidemment que
ces deux souches diffèrent pour plus d’un gène, c’est-à-dire au moins deux gènes.
Cela peut être évident quand les gènes étudiés gouvernent des couples de phénotypes
différents, car il apparaît en F2 des phénotypes eux-mêmes recombinés différant
des phénotypes parentaux (voir exercices).
Si les deux gènes étudiés sont impliqués dans un même phénotype, il n’y aura
toujours que deux phénotypes parentaux et aucun phénotypes recombinés, mais ce
sont les proportions de ces phénotypes en F2 qui mettront en évidence la recombinaison
génétique et l’existence de ces deux gènes, par des proportions différant de la
ségrégation 2/2 et caractéristiques de celles attendues pour deux couples d’allèles
indépendants ou non (voir plus loin, remarque 2, et exercices).
Si les deux souches parentales étudiées diffèrent pour plus d’un gène, il convient
alors de voir si elles peuvent différer pour deux gènes seulement; en effet, dans un
tel cas, on doit observer une ségrégation 2/2 pour chacun des gènes (chacun des
couples de phénotypes) pris isolément.
L’analyse génétique consiste alors à reconstruire, à partir des observations, la
méiose du diploïde F1 en spécifiant le contenu génétique des différents gamètes
produits et leurs fréquences respectives afin de « tester l’indépendance génétique »
en répondant à la question : les gamètes recombinés sont-ils équifréquents aux
gamètes parentaux ou non ?
• Si la réponse est oui, les deux gènes étudiés sont génétiquement indépendants, ce
qui conduit (sauf dans des cas exceptionnels, voir analyse de tétrades et exercices) à
la conclusion que les deux gènes sont, soit physiquement indépendants soit physiquement
liés, à une distance assez grande pour que la ségrégation des allèles au
premier locus soit indépendante de la ségrégation des allèles au second.
• Si la réponse est non, c’est que la fréquence des gamètes recombinés est inférieure
à celle des gamètes parentaux et qu’il y a liaison génétique.
Celle-ci est la conséquence
mécanique d’une liaison physique, avec une distance telle entre les locus,
qu’une fraction (1 – f ) des méioses se déroule sans crossing-over conduisant à
l’excès observé de gamètes parentaux.
On peut alors estimer une distance génétique
entre les locus des deux gènes.
En pratique, on fait le test de l’indépendance génétique en comparant les fréquences
des différents phénotypes parentaux ou recombinés observés aux fréquences attendues
de ces mêmes phénotypes, sous l’hypothèse d’indépendance génétique.
Bien évidemment, ces fréquences attendues ne sont pas les mêmes selon que le
double hétérozygote F1, dont on étudie la méiose, est croisé avec un autre F1 (croisement
F1 × F1) ou avec un parent récessif (test cross), pour les deux gènes étudiés.
Par ailleurs, il faut tenir compte, dans la reconstruction des phénotypes attendus et
le calcul de leurs fréquences respectives, des relations de dominance et de
récessivité; il suffit de rappeler que la ségrégation 2/2 se traduit par des proportions
de 3/4 de phénotypes dominants et 1/4 de phénotypes récessifs dans un croisement
F1 × F1 et par 1/2 de phénotypes dominants et 1/2 de phénotypes récessifs dans un
croisement F1 × parent récessif (test cross).
Qu’attend-on concrètement dans le cas de deux gènes ?
B - Test de l’indépendance génétique à l’issue d’un croisement F1 × F1
Si on croise une souche pure (A//A; B//B), de phénotype [A, B], par une souche pure
(a//a; b//b), de phénotype [a, b], l’hétérozygote F1 sera de génotype (A//a; B//b) et
de phénotype [A, B], s’il y a dominance des deux phénotypes parentaux [A] et [B].
La méiose, chez un individu F1 peut alors produire quatre types de gamètes : deux
gamètes de type parental (A, B) et (a, b) et deux gamètes de type recombiné (A, b) et
(a, B).
Si on note r, la fréquence des gamètes recombinés, chacun des deux types de
gamètes recombinés aura une fréquence égale à r/2, et chacun des deux types de
gamètes parentaux aura une fréquence égale à (1 – r)/2.
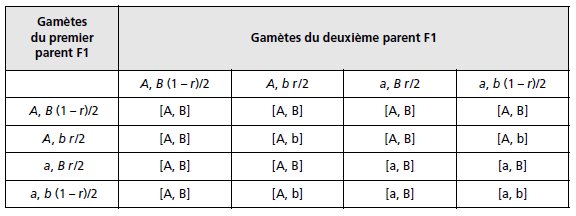
TABLEAU 3.1 TABLEAU DE CROISEMENT DES GAMÈTES FORMÉS À LA MÉIOSE POUR DEUX COUPLES D’ALLÈLES.
r étant la fréquence des gamètes recombinés; les phénotypes [A] et [B] étant
supposés dominants.
Dans le cas d’un croisement F1 × F1, les résultats attendus peuvent être formulés
dans un tableau de croisement des gamètes (tab. 3.1), où les phénotypes des
diploïdes résultant de l’union des gamètes dépendront des relations de dominance et
de récessivité existant pour chacun des couples d’allèles.
Les fréquences attendues des quatre types de phénotypes résultant des unions des
quatre types de gamètes sont égales à :
f [A, B] = (3 – 2r + r2)/4; f [A, b] = f [a, B] = r(2 – r)/4; f [a, b] = (1 – r)2/4
Si les deux gènes sont génétiquement indépendants, r = 1/2, ce qui correspond
aussi à (1 – r) = 1/2, équifréquence des gamètes recombinés et parentaux.
On
retrouve alors les proportions 9/16-3/16-3/16-1/16, encore notées 9-3-3-1, déjà
observées par Mendel.
S’il y a liaison génétique, alors r est inférieur à 1/2, et la proportion des phénotypes
parentaux [a, b] doit être significativement supérieure à 1/16, ce qui doit éventuellement
être justifié par un test statistique.
Cette fréquence tend vers 1/4 à mesure
que la distance diminue.
Remarque 1. La distance est facile à estimer par l’estimation de :
r = 1 – 2√f[a,
b]
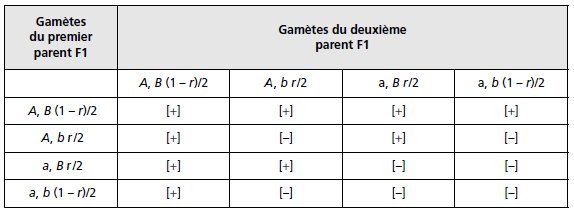
Remarque 2. Si les deux gènes sont impliqués dans un même phénotype
mutant, le tableau de croisement des gamètes (tabl. 3.2) ne laissera apparaître
que les deux seuls phénotypes parentaux sauvage [+] ou mutant [–].
TABLEAU 3.2 TABLEAU DE CROISEMENT DES GAMÈTES, GÉNOTYPES ET PHÉNOTYPES ASSOCIÉS.
Lorsque deux gènes ont des actions complémentaires sur un même phénotype,
sauvage s’il y a au moins un allèle A et un allèle B, mutant si A ou/et B sont
absents. r étant le taux de recombinaison entre les deux gènes.
Les fréquences attendues des deux types de phénotypes résultant des unions des
quatre types de gamètes sont égales à :
f [+] = (3 – 2r + r2)/4; f [–] = (1 + 2r – r2)/4
Estimer r revient à résoudre l’une des deux équations précédentes.
On observe, dans le cas de deux gènes indépendants, avec l’équifréquence des
gamètes, c’est-à-dire r = 1/2, 9/16 de phénotypes sauvages et 7/16 de phénotypes
mutés, où la proportion 7/16 est la somme 3/16 + 3/16 + 1/16 correspondant aux
trois types de phénotypes mutés quand les deux gènes (cas précédent) gouvernent
des caractères différents.
Selon le type d’interaction entre deux gènes indépendants affectant un même
caractère, on peut observer des regroupements partiels différents des quatre proportions
9-3-3-1 (voir exercices).
Remarque 3. À la limite, si deux gènes impliqués dans deux phénotypes
distincts (tab. 3.1) sont si proches que leur distance est nulle et qu’aucun
crossing-over ne survient, les allèles A et B vont coségréger et on retrouvera
3/4 de phénotype parental [A, B] dominant, et 1/4 de phénotype
parental récessif [a, b] (il suffit de prendre r = 0 dans les équations précédentes).
Un tel résultat pourrait alors être interprété comme une ségrégation 2/2 pour
un couple d’allèle et le phénotype parental [a, b] pourrait être formellement
interprété comme résultant d’une mutation unique et pléiotrope, responsable
simultanément des deux phénotypes [a] et [b].
Dans le cas où les deux mutations a et b conduisent à un même phénotype
mutant [–] (tab. 3.2), l’absence de gamètes recombinés, si les deux gènes sont
très liés, conduirait à 3/4 [+] et 1/4 [–].
Ce résultat serait aussi interprétable
comme résultant d’une ségrégation 2/2, conduisant à la conclusion que les
souches mutées et sauvages ne diffèrent que pour un seul gène.
C’est pourquoi l’interprétation d’une ségrégation 2/2 est souvent complétée
d’une remarque de précaution précisant que les souches mutées et sauvages
peuvent éventuellement différer pour deux gènes suffisamment proches pour
que le nombre de méioses étudiées en F1 n’ait éventuellement pas permis
d’observer une seule recombinaison par crossing-over, et donc un écart significatif
à la ségrégation 2/2.
Remarque 4. Bien évidemment, si l’un des couples d’allèles (ou les deux)
gouverne(nt) des phénotypes codominants, le problème est en fait plus
simple, puisqu’il existe une correspondance bi-univoque entre phénotype et
génotype qui permet de déterminer sans ambiguïté le contenu génétique de
chacun des gamètes, pour ce(s) gène(s).