On distingue, chez les chiens de la race Labrador, trois types de phénotypes
de robe (couleur du pelage), la robe noire, la robe dorée et la robe
chocolat.
On dispose de trois souches pures pour chacune de ces robes et
on se propose d’étudier le déterminisme génétique de ces phénotypes.
1. La souche A [noire] est croisée avec la souche B [chocolat], les descendants
F1 sont tous de couleur noire.
Les croisements F1 × F1 donnent 3/4 [noire] et 1/4 [chocolat]. Que peuton
en conclure ? Par quel autre type de croisement aurait-on pu arriver à la
même conclusion ? Quels auraient été les résultats observés pour ce type
de croisement ?
2. La souche B [chocolat] est croisée avec la souche C [dorée], les descendants
F1 sont tous de couleur noire.
Le croisement d’un F1 par un parent [dorée] donne 50 % de [noire]
et 50 % de [dorée], mais les croisements F1 × F1 donnent 9/16 [noire],
3/16 [chocolat] et 4/16 [dorée].
Concluez, en montrant que ces résultats sont cohérents avec ceux de la
question précédente.
3. Quels seraient les résultats et l’interprétation génétique de l’analyse des
croisements entre les souches [noire] et [dorée] suivis par l’étude F1 × F1
ou test cross ?
➤ Niveau Licence /Définition des objectifs.
– Modification des proportions classiques de ségrégation dans le cas d’interaction
entre gènes indépendants.
– Exemple de
la race de chiens labrador.
Solution
1. On a une ségrégation 2 × 2 typique d’un couple d’allèles qu’on notera A et a; la souche
[noire] étant homozygote A//A et la souche [chocolat] étant homozygote a//a. Le phénotype
[chocolat] est récessif devant le phénotype [noir].
Un test cross F1 × parent chocolat aurait
donné 50 % de descendants [noir] et 50 % de [chocolat].
2. Les résultats de ce croisement sont inhabituels à plusieurs titres :
– le phénotype de la F1 n’est pas celui d’un des deux parents;
– le test cross, même s’il fournit des proportions 1/2-1/2, ne présente pas la réapparition du
phénotype parental [chocolat] qu’on devrait observer si les souches parentales différaient
pour un seul gène;
– le croisement F1 × F1 permet la réapparition de ce phénotype parental mais ne présente
pas les rapports 3/4-1/4 typiques d’une ségrégation 2/2; au contraire, on observe les
proportions modifiées d’une ségrégation 9-3-3-1 pour deux gènes indépendants, comme si
deux classes de descendants, de fréquence 3/16 et 1/16, présentaient un même phénotype
[dorée], d’où une proportion de 4/16.
Il n’y a donc pas de ségrégation 2/2; les souches B et C diffèrent pour au moins deux gènes.
Comme on sait que les phénotypes [noire] et [chocolat] dépendent des allèles d’un même
gène, que tous les F1 du croisement étudié sont [noire] et que les souches sont pures (homozygotes),
on peut en déduire que la souche C est de génotype {A//A; c//c}, où c est l’allèle
d’un gène qui gouverne la robe [dorée], malgré la présence des allèles A sur le premier gène;
l’effet de l’allèle c est de bloquer l’expression de l’allèle A. Cet effet est récessif puisque les
F1 (A//a; C//c) sont de robe noire.
La souche A est de génotype {A//A; C//C} où l’allèle C du deuxième gène permet l’expression
de l’allèle A conduisant à la robe noire.
La souche B [chocolat] serait alors {a//a; C//C}.
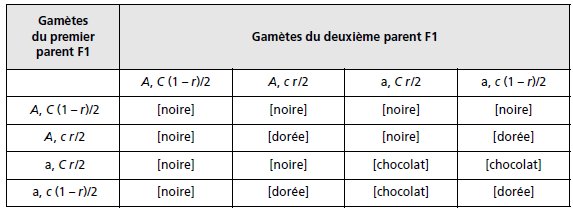
Le croisement de la souche B avec la souche C donne un double hétérozygote pour les deux
gènes, ce qui conduit à un tableau à quatre gamètes et aux proportions 9-3-3-1 si les gamètes
sont équifréquents (indépendance génétique des deux couples d’allèles), et à un regroupement
pour une proportion de 4/16 de tous les génotypes c//c qui seront de robe dorée, que le
génotype soit A//A ou A//a ou a//a (tabl. 3.25).
Il y a épistasie du deuxième gène sur le premier, avec un effet récessif de l’allèle épistatique.
Conclusion : Quand deux gènes sont indépendants et que l’un exerce un effet épistatique
récessif sur l’autre, on observe dans le croisement F1 × F1, une modification des proportions
9-3-3-1 en un regroupement 9-3-4.
TABLEAU 3.25 GÉNOTYPES ET PHÉNOTYPES DE LA F2
APRÈS UNION DES GAMÈTES ISSUS DE LA MÉIOSE CHEZ LA F1.
3. Le croisement des souches A [noire] de génotype (A//A; C//C) et C [dorée] (A//A; c//c)
donnera des descendants F1 de couleur noire.
Les croisements F1 × F1 donneront 3/4 [noire]
et 1/4 [dorée], car tous les individus sont homozygotes pour le gène A; et le test cross
donnera 50 % de [noire] et 50 % de [dorée].
On conclura, avec justesse, qu’il y a ségrégation 2/2, et que les souches A et C ne diffèrent
que pour un seul gène, où un allèle récessif c est responsable de la robe dorée, et un allèle
dominant C est responsable de la robe noire.
Rien ne peut laisser supposer que la couleur de la robe dépend en fait d’un autre gène qui est
sous la dépendance épistatique du gène étudié dans les croisements entre souches A [noire]
et C [dorée].
Ce sont les résultats inhabituels des croisements entre souches B [chocolat] et
C [dorée] qui permettent cette conclusion.
Exercice 3.9
:
On croise, chez Drosophila melanogaster, une souche pure sauvage de
référence (SSR), aux yeux [rouge brique], avec une souche A pure, présentant
également des yeux de couleur sauvage [rouge brique].
Les individus F1 ont des yeux [rouge brique], mais les croisements entre
F1 donnent chez les individus F2 :
– phénotypes [rouge brique] : 13/16;
– phénotypes [rouge vif] : 3/16.
Interprétez ces résultats et proposez un moyen expérimental permettant de
choisir entre les deux conclusions possibles.
➤ Niveau Licence (L3)/Définition des objectifs.
– Modification des proportions classiques de ségrégation dans le cas d’interaction
entre gènes indépendants.
– Exemple de la drosophile.
Solution. La simple observation des phénotypes F2, sachant que les souches parentales sont
pures, conduit d’abord à des conclusions fonctionnelles.
Les deux souches parentales ont le même phénotype sauvage mais ne sont certainement pas
toutes les deux de génotype sauvage.
En effet, on voit apparaître un phénotype mutant en F2,
ce qui prouve l’existence d’une mutation conférant la couleur rouge vif chez la souche parentale
A.
Cette mutation est à l’état homozygote puisqu’il s’agit d’une souche pure.
Comme la
souche A présente un phénotype sauvage, on doit en conclure qu’elle est aussi mutante dans
un gène appelé « suppresseur », dont l’allèle mutant « supprime » l’effet de l’allèle conférant
la couleur rouge vif, restaurant ainsi un phénotype sauvage [rouge brique].
Par ailleurs, sachant que les proportions observées constituent un rapport modifié de la suite
classique 9-3-3-1, les deux souches diffèrent pour deux gènes indépendants.
Comme il n’y a
pas de crossing-over chez le mâle de la drosophile, on peut même préciser que les deux gènes
en question sont physiquement indépendants.
Les souches étant pures et homozygotes, on peut écrire ainsi leurs génotypes :
– souche SSR : {a+//a+; su+//su+};
– souche B : {a//a; su//su}.
Remarque.
On ne peut, pour l’instant, définir les relations de dominance et de récessivité,
et les allèles sont notés « + » quand il désigne l’allèle de la souche SSR,
su étant l’allèle ayant un effet suppresseur sur l’allèle a conférant la couleur rouge vif.
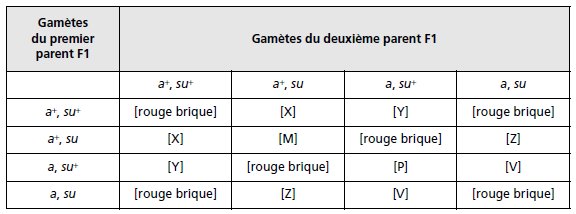
Les descendants F1 auront pour génotypes {a+//a; su+//su}, ce qui donnera, à l’issue du croisement
F1 × F1, les divers génotypes (tabl. 3.26) dont certains ont un phénotype connu.
TABLEAU 3.26 GÉNOTYPES ET PHÉNOTYPES DE LA F2
APRÈS UNION DES GAMÈTES ISSUS DE LA MÉIOSE CHEZ LA F1.
Connaissant le phénotype des souches parentales et de la F1, il est facile de compléter le
tableau pour les génotypes correspondants.
Il reste à définir les phénotypes [M], [P], [X],
[Y], [Z] et [V] relatifs aux nouveaux génotypes générés par la recombinaison génétique, en
fonction de l’effet de dominance ou de récessivité des allèles mutés, a et su, face aux allèles
sauvages a+ et su+ (tabl. 3.27).
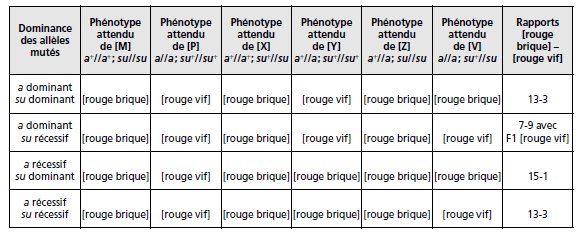
Si l’allèle suppresseur su a un effet récessif, il n’y aura suppression que chez les homozygotes
su//su, et l’allèle su n’aura aucun effet chez les hétérozygotes su+//su. Au contraire,
si l’effet de l’allèle su est dominant il y aura un effet suppresseur aussi bien chez les su//su
que chez les su//su+.
TABLEAU 3.27 COMBINAISONS POSSIBLES DE DOMINANCE ET DE RÉCESSIVITÉ.
Pour les allèles mutants a et su vis-à-vis de leurs allèles sauvages respectifs a+ et su+,
détermination des six phénotypes [M], [P], [X], [Y], [Z] et [V] qui en résultent et du
rapport 9-3-3-1 modifié qui en résulte.
Le fait que la F1, de génotype su//su+, soit de phénotype [rouge brique] ne nous permet pas
de trancher, car on ne sait pas si l’allèle a est récessif ou dominant vis-à-vis de a+.
Dans le
premier cas (a récessif), peu importe que su soit récessif ou dominant, puisque de toute façon
le phénotype [rouge brique] ne dépend que de la seule présence de A; dans le deuxième cas
(a dominant), cela prouverait que su l’est aussi, puisqu’alors l’effet dominant de a devrait
être corrigé par l’effet dominant de su, le F1 étant su+//su.
Le rapport observé 13-3 est compatible avec l’hypothèse que les allèles mutés sont tous deux
récessifs ou tous deux dominants.
Remarque 1. Le deuxième cas (a dominant/su récessif) conduirait à une F1 [rouge
vif], ce qui donnerait en F2 un rapport 7-9.
Ce type de rapport a déjà été vu dans le cas
où deux gènes ont des effets complémentaires et où le phénotype sauvage suppose la
présence d’au moins un allèle sauvage de chaque gène, mais on a alors 9 sauvages
pour 7 mutants, alors qu’ici, dans le cas d’une suppression récessive d’une mutation
dominante, on a 9 mutants pour 7 sauvages.
Remarque 2. Le troisième cas (a récessif/su dominant) conduit à un rapport 15-1 déjà
vu dans le cas de deux gènes redondants (voir exercice 3.6, sur la forme des fruits de
la bourse à pasteur), or ce cas est formellement identique si on veut bien considérer
que l’allèle sauvage d’un gène dupliqué joue le rôle de suppresseur dominant de tout
allèle muté récessif de l’autre gène !
Conclusion : Quand deux gènes sont indépendants et que la mutation récessive de l’un voit
son effet « supprimé » par la mutation récessive de l’autre, on observe dans le croisement
F1 × F1, une modification des proportions 9-3-3-1 en un regroupement 13-3.
Il en est de
même si les mutations des deux gènes sont toutes deux dominantes.
Pour choisir entre les deux interprétations possibles du rapport 13-3, il suffit de faire un test
cross entre les F2 [rouge vif] et un parent A apportant un gamète (a; su).
• Si les mutations a et su sont dominantes, tous les descendants F3 seront [rouge brique],
puisqu’ayant reçu, de toute façon, a et su du parent A.
• Si les mutations a et su sont récessives, un tiers des descendants sera de phénotype [rouge
vif].
En effet, 2/3 des F2 testés sont de génotype {a//a; su+//su} et la moitié de leur gamètes
(a; su+), en s’unissant aux gamètes (a; su), donneront des génotypes {a//a; su+//su}, de
phénotype [rouge vif] si la mutation su est récessive.