La drosophile, Drosophila melanogaster, est un organisme diplobiontique (phase
haploïde réduite aux gamètes). La fécondation des ovocytes est réalisée au cours de
la ponte (jour j) à partir d’un stock de spermatozoïdes conservés dans une spermathèque
à l’issue de la copulation.
Durant le premier jour, les divisions conduisent à la formation d’un blastoderme,
au sein duquel la lignée germinale s’est isolée, puis les mouvements cellulaires
(gastrulation) aboutissent à la formation des trois feuillets, ectoderme, endoderme et
mésoderme, à la métamérisation de l’embryon (segmentation préfigurant les segments
tête-thorax-abdomen de l’adulte) et à l’éclosion du premier stade larvaire (j + 1).
À l’issue de deux mues successives, le troisième stade larvaire forme une
pupe (j + 5) au sein de laquelle se constitue l’imago adulte par apoptose des tissus
larvaires et morphogenèse des tissus adultes à partir de massifs de cellules (disques
imaginaux) demeurées dans un état prédifférencié.
L’émergence de l’adulte survient à j + 10; les femelles restent vierges pendant
quelques heures avant de pouvoir s’accoupler, ce qui permet de les recueillir et de les
isoler afin de réaliser les croisements expérimentaux avec les mâles choisis.
La
longueur du cycle vital dépend de la température, les valeurs données ici correspondent
à 25 °C.
On rappelle que le génome de drosophile est réparti sur cinq chromosomes, les
autosomes 2, 3 et 4, le chromosome 1 (ou X) et le Y.
Le sexe est hétérogamétique,
les femelles étant XX et les mâles XY, mais, en réalité, le sexe est déterminé par le dosage entre le lot d’autosomes et le lot de X.
Il reste que le sexe mâle est, sauf
exception, hémizygote pour les gènes de l’X.
Il n’y a pas de crossing-over à la
méiose chez le mâle.
Problèmes
:
Problème 11.1
Un organisme transgénique est un organisme dont le génome a été transformé
par l’insertion (ou l’addition) d’un vecteur moléculaire porteur
d’une information génétique additionnelle.
Pour la bactérie, les vecteurs parmi les plus utilisés sont des plasmides des
transposons ou des dérivés du virus λ.
Chez Drosophila melanogaster on
utilise les « éléments P », des séquences transposables d’ADN capables de
s’insérer plus ou moins aléatoirement dans l’ADN génomique, selon un
mécanisme différent de celui des rétrovirus.
En général, on utilise un
élément P déficient, capable de s’insérer mais incapable, après son insertion,
de transposer (excision suivie d’une réinsertion en un autre point du
génome, ou duplication suivie d’une insertion de la copie en un autre point
du génome).
Le protocole de base consiste à injecter une quantité d’éléments P, porteurs
d’un transgène d’intérêt, dans des oeufs de drosophile, juste après la ponte,
afin de récupérer à la pupaison des adultes F0.
L’organisme F0 est une
mosaïque cellulaire et, seule, une fraction variable des tissus somatique et
germinal sera constituée de cellules transgéniques, ayant été génétiquement
transformées par l’insertion d’un élément P (un seul par cellule car le
phénomène est assez rare).
On croise F0 avec un parent choisi afin de récupérer, dans les quelques cas
où F0 a fourni un gamète transgénique, des individus F1 porteurs de
l’élément P et du transgène dans toutes ses cellules.
Il est crucial, à ce
stade du protocole, de pouvoir distinguer les F1 porteurs du transgène et
ceux qui en sont dépourvus (voir question 1).
Par croisement entre F1, on peut alors obtenir 1/4 d’individus F2 homozygotes
pour le transgène.
Dans le problème qui suit on considérera que les drosophiles transgéniques
F1 étudiées sont porteuses d’un seul élément P, inséré au hasard en un
seul endroit de leur génome.
On dispose d’une souche pure A de drosophile, homozygote pour une
mutation de perte de fonction (notée adh0) dans le gène de l’alcooldéshydrogénase
(localisé sur le chromosome 2).
Cette déficience en alcool-déshydrogénase entraîne l’incapacité d’oxyder
totalement les alcools (éthanol, propanol, butanol) et rend l’homozygote adh0//adh0 sensible à la présence, dans le milieu nutritif, de
penténol (celui-ci est partiellement dégradé en une cétone très toxique pour
l’organisme).
Ce phénotype de sensibilité au penténol est récessif comme
la déficience enzymatique.
Au sein de la souche A, on obtient deux mâles F1 transgéniques m1 et m2,
résistants au penténol, par insertion, dans leur génome, d’un élément Pa
porteur de la séquence totale du gène de l’alcool-déshydrogénase (notée adh+).
On entreprend leur analyse génétique.
1. Quel avantage peut-on tirer de ce retour à la résistance au penténol pour
améliorer l’efficacité du protocole de construction de drosophiles transgéniques,
rappelé plus haut, pour un autre transgène que celui de l’alcooldéshydrogénase
?
2. Le mâle transgénique m1 est croisé avec une femelle de la souche A, on
obtient des femelles, toutes résistantes au penténol, et des mâles, tous
sensibles au penténol.
Le mâle transgénique m2 est croisé avec une femelle A.
La moitié des
femelles et des mâles sont résistants au penténol.
Ces observations permettent-elles d’assigner, chez m1 et m2, l’insertion de
l’élément P et du transgène sur un chromosome précis ?
Vous ferez un schéma clair des chromosomes parentaux du mâle m1 et de
la femelle A.
3. Le mâle transgénique m2 est croisé avec une femelle B de souche pure
pour la mutation adh0 et pour la mutation « cardinal » (qui sera nommée cd)
du chromosome 3, conférant un phénotype récessif [oeil pourpre].
La moitié des femelles F2 et la moitié des mâles F2 sont résistants au penténol.
Un mâle F2 résistant au penténol est croisé en test-cross avec une
femelle B; on obtient dans chaque sexe deux phénotypes équifréquents
[oeil pourpre, sensibles au penténol] et [oeil sauvage, résistantes au
penténol].
Interprétez ce résultat en décrivant par un schéma clair les génotypes
(et les chromosomes) impliqués dans ces croisements.
4. Le mâle transgénique m2 est croisé avec une femelle C de souche pure
pour la mutation adh0 et pour la mutation « vestigiale » (qui sera nommée vg)
du chromosome 2, conférant un phénotype récessif [aile vestigiale].
La
moitié des femelles F2 et la moitié des mâles F2 sont résistants au penténol.
Un mâle F2 résistant au penténol est croisé en test-cross avec une femelle C;
on obtient dans chaque sexe quatre phénotypes équifréquents, [aile vestigiale,
sensibles au penténol], [aile vestigiale, résistantes au penténol], [aile
normale, sensibles au penténol] et [aile normale, résistantes au penténol].
Interprétez ce résultat en décrivant par un schéma clair les génotypes
(et les chromosomes) impliqués dans ces croisements et montrez en quoi il
est cohérent avec le résultat de la question précédente.
5. Le test-cross de la question 3 est réalisé entre une femelle F1 résistante
au penténol et un mâle de la souche B.
On obtient dans chacun des sexes
quatre phénotypes équifréquents, [oeil pourpre, sensibles au penténol], [oeil
pourpre, résistantes au penténol], [oeil sauvage, sensibles au penténol], [oeil
sauvage, résistantes au penténol]. Qu’en concluez-vous ?
Solution
1. Les organismes de la souche A sont adh0//adh0 (sur leur chromosome 2) et présentent le
phénotype de sensibilité au penténol qui sera noté [penS].
Les mâles transgéniques m1 et m2
sont logiquement [penR] puisqu’ils sont porteurs, par l’insertion d’un élément P, du transgène
adh+.
La résistance au penténol peut être très utile car elle peut servir à la fois de crible de sélection
positive et de test d’identification phénotypique des F1 porteurs d’un élément P.
Il suffit, en
effet, dans le protocole rappelé plus haut, de croiser les F0 avec des drosophiles de la souche A
et de les faire pondre sur un milieu additionné de penténol; seuls survivent les F1 résistants
c’est-à-dire génétiquement transformés par un élément P.
Le transgène adh+ peut ainsi servir de marqueur de sélection des transformés comme les
gènes de résistance aux antibiotiques pour les plasmides bactériens ou les gènes URA3 ou
LYS2 pour les plasmides de levure, le transgène d’intérêt étant alors une autre séquence
clonée dans le même élément P.
2. Les drosophiles transgéniques m1 et m2 étudiées sont porteuses d’un seul élément P (ce
sont des F1), inséré au hasard en un seul endroit de leur génome.
La question posée est
d’assigner l’insertion à un chromosome chez m1 et m2, sachant que la localisation de cette
insertion est sans doute différente chez m1 et m2 (insertion au hasard).
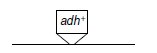
On différenciera le transgène dans l’écriture des génotypes, en figurant son insertion de la
manière suivante sur le chromosome porteur :
Deux types d’insertion sont possibles et décrits par les figures 11.1 et 11.2.
Si le transgène est inséré sur le chromosome X (fig. 11.1), celui-ci est transmis à toutes les
femelles F2 qui seront toutes résistantes au penténol et à aucuns mâles qui seront tous sensibles.
Ce cas correspond au mâle m1.
Si le transgène est inséré sur un autosome A (fig. 11.2), cet autosome A étant transmis à un
descendant sur deux, indépendamment du sexe, la moitié des femelles, comme la moitié
des mâles, seront résistants, et les deux autres moitiés sensibles.
Ce cas correspond au
mâle m2.
Figure 11.2 Génotype du mâle
transgénique avec insertion de l’élément P
sur un autosome et génotype de la femelle A croisée avec lui.
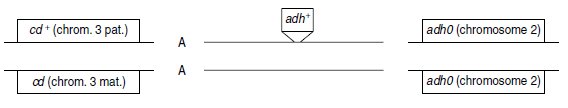
3. Le transgène du mâle m2 est porté par un autosome; le mâle F2 résistant, issu du croisement
avec B, sera lui-même porteur du transgène paternel, et aura le génotype suivant :
où A est un autosome qui peut être le chromosome 2 ou 3 ou 4.
Le test-cross avec une femelle B permet de montrer que le phénotype [oeil sauvage] coségrège
avec le phénotype [penR].
Le phénotype [oeil sauvage] est dû à la transmission de l’allèle paternel cd+, tandis que le
phénotype [penR] est dû à la transmission du transgène adh+, lui-même, sur un autosome
paternel.
La cotransmission systématique de cd+ et adh+ conduit, sachant qu’il n’y a pas de crossingover
chez la drosophile mâle, à la conclusion que les deux séquences cd+ et adh+ sont physiquement
liées (sinon on pourrait avoir des gamètes recombinés transmettant cd et adh+) et à
la conclusion que le transgène adh+ a été inséré sur le chromosome 3 du mâle m2.
Remarque. Peu importe la distance entre le locus de cd et le site d’insertion du transgène
puisqu’il n’y a pas de crossing-over chez le mâle drosophile, ce qui explique la
coségrégation absolue.
4. On sait que le transgène est inséré sur le chromosome 3 et qu’il est, de ce fait, physiquement
indépendant du locus vg, situé sur le chromosome 2.
Il est donc logique d’observer une
ségrégation indépendante des deux caractères avec 25 % de chacun des phénotypes possibles.
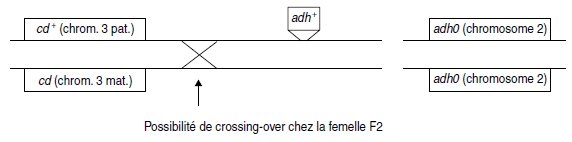
5. Evidemment, dans ce cas, on teste chez la femelle résistante, la recombinaison par
crossing-over à la méiose, afin de mesurer la liaison entre le site d’insertion du transgène et
le locus cd, sur le chromosome 3 issu du mâle m2. En effet le génotype de cette femelle résistante
s’écrit :
La ségrégation indépendante des caractères phénotypiques malgré la liaison physique entre
leurs déterminants génétiques (le couple d’allèles cd et cd+ et le transgène adh+, sans correspondant
sur l’autre autosome) montre que le locus cd et le site d’insertion du transgène sont
génétiquement indépendants, c’est-à-dire qu’ils sont suffisamment éloignés pour que le taux
de recombinaison soit égal à 50 %.