4. a Chacune des quatre souches H, I, J et K est porteuse d’un chromosome balanceur
pistable par la mutation bar et d’un chromosome muté qu’on peut appeler h, i, j ou k.
Les femelles de ces quatre souches ont respectivement les génotypes FM3//i, FM3//j, FM3//h,
FM3//k.
Le fait que les mutations portées par les chromosomes h, i, j et k soient maintenus en stock
face au balanceur, prouve qu’elles ont toutes un effet récessif vis-à-vis de leurs homologues
respectifs sur FM3.
Le croisement de femelles H, I, J ou K avec des mâles sta-1/Y donne des femelles F1.
Celles
qui sont de phénotype [B+] n’ont pas reçu le balanceur et sont donc de génotype sta-1//h,
sta-1//i, sta-1//j ou sta-1//k.
On identifie ainsi les individus dont le génotype est porteur de
mutations sur les deux chromosomes X.
Si l’un de ces génotypes, sta-1//i, par exemple, s’avère stérile, on peut conclure que le mutant I
est affecté dans le gène STA (pas de complémentation fonctionnelle).
Ce test de complémentation fonctionnelle permet de conclure que les quatre mutants H, I, J
et K sont mutés dans le gène STA.
NB :
En absence de test de ségrégation 2/2, on ne peut savoir si les mutants H, I, J ou K sont
des mutants simples, mutés dans le seul gène STA (ce qui est sûr, compte tenu du TCF) ou si
ce sont des mutants multiples touchés aussi dans un autre gène.
4. b Les mâles H sont h//Y; les femelles H sont FM3//h. Leur croisement donnent deux types
de femelles F1, FM3//h de phénotype [B/2] et h//h de phénotype [B+].
Ces dernières ont toujours des ailes courtes, ce qui signifie que le phénotype [aile courte] est
récessif et associé à la « mutation h ».
De la même manière le phénotype [abdomen déformé] est récessif et associé à la « mutation
k ».
Cependant, il convient d’écrire « mutation h » ou « mutation k », car, comme cela a été dit,
on ne sait si les mutants étudiés sont simples ou multiples. Plusieurs hypothèses différentes
peuvent rendre compte de ces phénotypes et aussi de la ségrégation 2/2 observée.
– Hypothèse 1. Le mutant H (ou K) est simple.
Dans ce cas, le seul gène muté est STA (TCF)
et la mutation a un effet pléiotrope se traduisant à la fois par la stérilité femelle et un caractère
morphologique (aile courte ou abdomen déformé). La ségrégation 2/2 est alors
attendue.
– Hypothèse 2.
Le mutant H (ou K) est double.
Il est à la fois muté dans le gène STA et aussi,
indépendamment, dans un autre gène (du chromosome X) qui conditionne la morphogenèse
des ailes ou de l’abdomen.
La ségrégation 2/2 est logique (pas de crossing-over
chez le mâle, ni, ici, chez la femelle, en raison du chromosome balanceur).
Cette hypothèse est peu vraisemblable car la probabilité d’avoir un double mutant est égale
au carré de la probabilité d’en avoir un simple.
Si la probabilité d’un simple mutant spontané
est de l’ordre de 10–6 à 10–7, celle d’un double mutant devient de l’ordre de 10–12 à 10–14
mutants, c’est-à-dire impossible.
Il est vrai que l’induction de mutants augmente considérablement l’ordre de grandeur des
taux, passant à 10–4 ou 10–5, ce qui rend l’hypothèse 2 possible, bien que toujours peu
probable.
– Hypothèse 3.
Le mutant H (ou K) est simple en ce sens qu’il est porteur d’une seule mutation,
mais deux gènes sont touchés simultanément, le gène STA et un autre impliqué dans
la morphogenèse de l’aile ou de l’abdomen.
Il s’agit dans ce cas d’une délétion (un seul
événement mutationnel) couvrant ces deux gènes.
La ségrégation 2/2 est attendue.
Cette hypothèse a l’avantage de combiner les deux précédentes en prenant la vraisemblance
statistique de la première (un seul événement mutationnel) et la vraisemblance biologique de
la seconde (deux fonctions touchées).
Il est alors nécessaire de définir un protocole de choix entre les hypothèses, test de ségrégation
2/2 ou autre test, éventuellement moléculaire.
5. a Des femelles FM3//h sont croisées par des mâles SSR, notés h+//Y. Les femelles F1 de
phénotype [B+] sont de génotype h//h+.
NB : Il est préférable de noter, dans ce croisement, les mâles sauvages par h+//Y et non sta+//Y,
car on ne sait pas si h se réduit au gène STA ou s’il recouvre plusieurs gènes.
Si on notait les
mâles par sta+//Y, les femelles issues du croisement seraient notées h//sta+ ce qui
tendrait à réduire h au seul statut d’allèle de STA (ce qu’il est de toute façon, mais peut-être
pas uniquement).
On observe, dans ce test cross, une ségrégation 2/2 typique avec deux types de mâles équifréquents
h//Y et h+//Y, et deux types de femelles équifréquentes h//h et h//h+.
Cette observation ne permet de choisir aucune des hypothèses :
– si l’hypothèse 1 est la bonne, on attend une ségrégation 2/2;
– si l’hypothèse 2 est la bonne, on peut avoir une ségrégation 2/2 si les deux gènes sont très
proches, ce qui est une condition supplémentaire rendant cette hypothèse encore moins
probable, mais on n’a pas démontré qu’elle était fausse;
– si l’hypothèse 3 est la bonne, on s’attend aussi à une ségrégation 2/2, comme le montre le
schéma ci-dessous.
À la méiose, l’appariement de deux chromatides non-soeurs, dont l’une est délétée pour les
deux gènes A et B, induira une boucle de non-appariement (appelée « boucle de délétion »)
sur l’autre chromatide.
La première chromatide est A– et B–, la seconde est A+ et B+, mais aucun crossing-over ne peut
générer des chromatides recombinantes A+ et B– ou A– et B+, car cela supposerait un crossingover entre les deux gènes en un point qui existe sur la chromatide (A+, B+) mais qui n’existe
plus sur la chromatide homologue délétée.
5. b Le gène miniature en 10E2, gouvernant par ses mutations un phénotype ailes courtes, est
dans la zone 10D8-10E4 où le gène STA a été assigné; il est logique de penser que le mutant
h pourrait être muté dans STA et dans m.
Le croisement réalisé est un TCF pour le gène m. Les femelles F1 obtenues ont le génotype
« h/m » et le phénotype [ailes courtes], ce qui prouve que la souche H, mutée dans STA,
est aussi mutée dans le gène miniature.
L’hypothèse 1 (mutant H simple et mutation pléiotrope) est donc exclue, et restent les deux
autres.
6. Les femelles F1, de génotype k//k+, sont croisées, en test-cross, avec des mâles k/Y.
Le fait qu’il y ait trois types de descendants femelles en F2 exclut la ségrégation 2/2 d’un
couple d’allèles à la méiose chez les femelles F1 k//k+.
On doit donc conclure que les mutants K
sont porteurs d’au moins deux mutations différentes sur le chromosome X, recombinables
par crossing-over.
L’une des mutations affecte le gène STA (voir l’absence de complémentation dans les croisements
femelles K × mâles sta-1/Y), l’autre mutation touche un gène impliqué dans la
morphogenèse de l’abdomen, noté ab.
Sous cette hypothèse minimale de deux gènes touchés, le génotype de la femelle F1 peut
s’écrire (sta–, ab–)//(sta+, ab+), ce qui conduit au tableau de croisement des gamètes
(tabl. 11.9).
On observe 277 et 321 individus femelles de phénotype parental et 63 individus d’un phénotype
non-parental, donc recombiné [abdomen normal, pondant des oeufs qui ne se développent
pas].
Quel est son génotype ?
Comme on sait que le phénotype abdomen déformé est récessif, il s’agit obligatoirement du
génotype recombiné ab–//ab+, donc du génotype (ab–; sta–)//(ab+; sta–), et il est logique qu’il
ponde des oeufs qui ne se développent pas, puisqu’il est sta–//sta–.
Le fait que l’homozygote k//k, c’est-à-dire (sta–, ab–)//(sta–, ab–) ne ponde pas d’oeufs est dû
au fait qu’il a l’abdomen déformé, phénotype épistatique sur celui associé au gène STA, si bien
que les génotypes (sta+//ab–)//(sta–; ab–) devraient aussi avoir le phénotype [abdomen déformé,
ne pondant pas d’oeuf], ce qui est confirmé par l’interprétation quantitative des résultats.
En effet, on sait que les génotypes parentaux sont équifréquents entre eux, de même les
génotypes recombinés.
Or ici 277 et 321 sont des valeurs significativement différentes
comme 63 et 0.
S’il y a 63 génotypes d’un type recombiné, il doit y en avoir la même quantité
de l’autre.
S’il n’y a que trois phénotypes au lieu de quatre, c’est que deux génotypes ont
un même phénotype.
Comme on a un phénotype recombiné et deux phénotypes parentaux,
c’est que l’un des génotypes recombinés a le même phénotype que le phénotype parental en
excès, soit [abdomen déformé, sans ponte], ce qui confirme bien ce qu’on attend.
Cela permet, par ailleurs, de calculer la fréquence de recombinaison, soit 63 × 2/641, d’où
une distance de 19,7 unités de recombinaison.
On voit qu’avec une mutagenèse aux rayons X on peut avoir des doubles mutants sans trop
de difficultés.
7. Une inversion entraîne une double coupure de l’ADN aux deux extrémités (appelées
pieds) de l’inversion.
Si le pied d’une inversion est situé dans la séquence codante d’un gène, celui-ci sera inactivé,
puisque l’instruction est détruite.
NB : Une inversion touchant un gène dans son promoteur peut mettre la séquence codante en
phase avec le promoteur d’un gène situé à proximité de l’autre pied et aboutir alors à une surtranscription s’il s’agit d’un promoteur fort.
Ici le mutant I, muté dans le gène STA, lui-même localisé entre 10D8 et 10E4, possède une
inversion entre 10E2 et 13B1; il est dès lors très vraisemblable pour ne pas dire certain que
le gène STA est touché par cette inversion ce qui précise sa localisation en 10E2, c’est-à-dire
très proche du gène miniature (question 5).
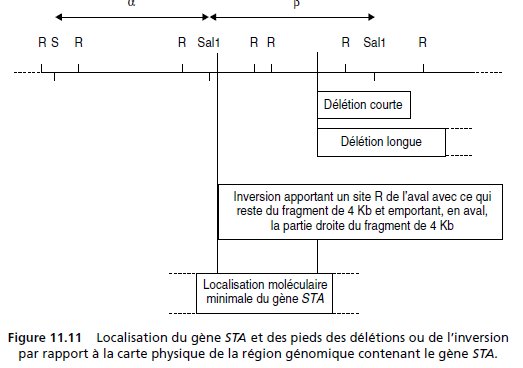
8. La mesure de la taille des fragments de restriction sur les southern blot pour la souche SSR
permet de compléter la carte de restriction (fig. 11.11).
Le fragment de 4 Kb est évidemment
reconnu par les sondes α et β.
L’absence de modification de la carte de restriction pour les mutants J et K plaide en faveur
de mutations ponctuelles du gène STA dans ces souches.
En revanche, les mutations affectant
le gène STA chez les mutants I et H sont associées à une modification de la carte de
restriction.
Ce résultat était attendu pour la souche I où l’analyse cytologique a montré une inversion, et
confirme l’hypothèse d’une délétion pour la souche H (questions 4 et 5.a).
Chez H la sonde α reconnaît les fragments présents chez la SSR; cette partie de la carte
physique n’est pas concernée par la délétion.
La sonde β identifie les fragments de 4 Kb et 1 Kb présents chez la SSR, mais identifie un
fragment de 5,5 Kb à la place de ceux de 5 et 6 Kb.
Deux solutions sont alors possibles pour situer l’ampleur de la délétion chez H :
– une délétion courte de 5,5 Kb autour de l’avant-dernier site R réduisant les deux fragments
de 5Kb et 6Kb à un fragment unique de 5,5 Kb (fig. 11.11);
– une délétion plus longue emportant les deux derniers sites R, et donc la totalité du fragment
de 6 Kb, raccordant la partie restante du fragment de 5 Kb (qui permet la reconnaissance
par la sonde β ) avec un site plus en aval et formant un nouveau fragment de
5,5 Kb.
L’analyse des fragments de restriction du mutant I permet de montrer que les fragments de 2,
7, 1, 5 et 6 Kb ne sont pas touchés, et que le seul fragment dont la taille est modifiée est le
fragment de 4 Kb, reconnu chez la SSR par les deux sondes.
L’un des pieds de l’inversion se situe dans ce fragment de 4 Kb :
– la sonde α reconnaît la partie droite du fragment qui est restée en place et se trouve soudée
avec l’autre pied de l’inversion qui apporte un site R, tel que le nouveau fragment fait 3 Kb
(fig. 11.11);
– la sonde β reconnaît la partie gauche de ce fragment qui, après l’inversion se retrouve
en 13B1 associée avec un site R en aval du pied de l’inversion, tel que le nouveau fragment
de restriction a une taille largement supérieure à 7 Kb (fig. 11.11);
– la sonde β reconnaît aussi la partie droite de ce fragment (3 Kb sur les deux southern), ce
qui prouve que le pied de l’inversion se situe à droite du site Sal1.
L’inversion du mutant I, qui touche le gène STA, prend pied dans le fragment de 4 Kb.
Dans
les deux hypothèses, le point de départ de la délétion du mutant H, qui touche le gène STA, se
situe dans le fragment de 5 Kb.
On peut donc conclure que la localisation moléculaire minimale du gène STA est de part et
d’autre du fragment EcoR1 de 1 Kb qui est totalement interne au gène STA.