On possède chez Drosophila melanogaster des mutants dans le gène STA
du chromosome X (mutation notée sta-1).
1. Les mâles sta-1/Y sont morphologiquement normaux. Croisés à des
femelles sauvages, ils donnent une descendance viable et fertile.
Les femelles sta-1//sta-1 sont morphologiquement normales, mais croisées
à des mâles sta-1/Y ou sta+/Y, elles pondent des oeufs qui ne se développent
pas.
Caractérisez l’effet phénotypique de la mutation sta-1 chez le mâle et la
femelle, en précisant à quel niveau du cycle vital de l’organisme s’exprime
le gène STA, après avoir défini les différents génotypes produits par ces
croisements et les conditions dans lesquelles ils sont produits.
2. Comme il n’est pas possible de maintenir en stock la mutation sta-1 au
sein d’une souche pure (les femelles sont stériles), la mutation sta-1 est
maintenue en stock face au balanceur FM3.
FM3 porte une mutation létale récessive et la mutation semi-dominante
Bar, responsable d’un phénotype [oeil réduit], noté [B/2], chez les femelles
hétérozygotes B/B+, ou [oeil très réduit], noté [B] chez les femelles homozygotes
B/B ou les mâles hémizygotes B/Y.
Le phénotype [oeil normal ou
sauvage] est noté [B+].
Vous établirez le protocole de croisement qui a permis d’aboutir à l’énoncé
des résultats de la question précédente.
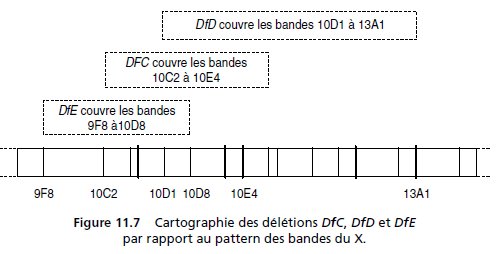
3. On dispose de trois souches porteuses d’une déficience (délétion)
partielle du chromosome X, notée DfC, DfD ou DfE, repérables sur les
chromosomes polyténiques des glandes salivaires (fig. 11.7).
Ces trois
déficiences sont conservées face au balanceur FM3.
On croise des mâles sta-1/Y par des femelles FM3/DfC (croisement 1),
FM3/DfD (croisement 2), ou FM3/DfE (croisement 3).
Les femelles F1 [B+] issues des croisements 1 et 2 sont, à leur tour, croisées
avec des mâles sauvages : elles pondent des oeufs qui ne se développent
pas.
Les femelles F1 [B+] issues du croisement 3 sont, à leur tour, croisées avec
des mâles sauvages : elles pondent des oeufs qui se développent en adultes.
Quelle est la localisation chromosomique du locus du gène STA ?
4. On obtient par mutagenèse de la souche SSR aux rayons X, quatre
nouvelles souches mutantes du chromosome X, notées H, I, J et K, dont les
mutations sont également conservées face au balanceur FM3.
a. Chacune des femelles H, I, J ou K, croisées avec un mâle sta-1/Y, donne
des femelles F1 de phénotype [B+] qui pondent des oeufs ne se développant pas.
On observe le même résultat quand on croise un mâle de chacune des
quatre souches par une femelle sta-1//sta-1.
Qu’en concluez-vous ?
b. Les mâles des souches I et J ont un phénotype morphologique tout à fait
sauvage; en revanche les mâles de la souche H ont des ailes courtes, et les
mâles de la souche K ont un abdomen déformé.
Mâles et femelles de la souche H donnent 50 % de femelles F1 de phénotype
[B/2, ailes sauvages] et 50 % de femelles F1 de phénotype [B+, ailes
courtes].
Mâles et femelles de la souche K donnent 50 % de femelles F1 de phénotype
[B/2, abdomen normal] et 50 % de femelles F1 de phénotype [B+,
abdomen déformé, ne pondant pas d’oeufs].
Enoncez les diverses hypothèses qui peuvent être faites a priori pour
rendre compte des résultats pour H et K, et discutez de la vraisemblance de
ces différentes hypothèses.
5. a. Le croisement des femelles H par des mâles sauvages donne 50 % de
femelles F1 de phénotype [B+] qui, croisées avec des mâles H, donnent
en F2 :
– femelles avec ailes normales, pondant des oeufs qui se développent en
adultes : 277;
– femelles avec ailes courtes, pondant des oeufs qui ne se développent
pas : 252;
– mâles avec ailes normales : 269;
– mâles avec ailes courtes : 257.
Ces résultats permettent-ils de choisir entre les hypothèses formulées
précédemment ?
b. On connaît l’existence d’un gène appelé miniature, (mutation notée m)
et localisé en 10E2 sur le chromosome X, dont les mutations m (à effet
récessif) entraîne un phénotype [ailes courtes].
Des femelles m/m sont croisées avec des mâles H; tous les descendants F1
ont des ailes courtes et les femelles pondent des oeufs qui se développent
en adulte.
Cela permet-il de choisir entre les hypothèses formulées sur la nature du
mutant H ?
6. Le croisement des femelles K par des mâles sauvages donne 50 % de
femelles F1 de phénotype sauvage qui, croisées avec des mâles K, donnent
en F2 :
– femelles avec abdomen normal, pondant des oeufs qui se développent en
adultes : 277;
– femelles avec abdomen normal, pondant des oeufs qui ne se développent
pas : 63;
– femelles avec abdomen déformé, ne pondant pas d’oeufs : 321;
– mâles avec abdomen normal : 327;
– mâles avec abdomen déformé : 315.
Ces résultats permettent-ils de choisir entre les hypothèses formulées sur la
nature du mutant K ?
Le génotype du mutant K sera précisé ainsi que le tableau de gamètes
conduisant aux résultats observés.
7. On croise des femelles FM3/sta-1, H, I, J et K par des mâles sauvages
puis on dissèque les glandes salivaires des larves F1 afin d’observer les
chromosomes polytènes.
On peut distinguer les larves femelles, ne possédant pas de chromosome Y
mais deux X; on peut aussi distinguer les femelles porteuses d’un chromosome
balanceur qui entraîne un grand nombre de boucles d’inversion.
Les caryotypes des larves femelles sans balanceur ne se distinguent pas
d’un caryotype sauvage pour toutes les larves F1 issues de femelles FM3/ sta-1, H, J ou K.
En revanche, le caryotype des larves femelles sans balanceur,
issu des femelles I présente une boucle d’inversion entre les bandes
10E2 et 13B1.
Qu’en concluez-vous sur la localisation cytologique du gène STA ?
8. L’ADN génomique correspondant à la région où est localisé le gène STA
a été cloné. On connaît la carte physique de cette région (fig. 11.8).
L’ADN génomique des souches sauvage, sta-1/sta-1, H, I, J et K est digéré
totalement par EcoR1; les fragments de restriction sont séparés par électrophorèse
sur gel d’agarose puis dénaturés et transférés sur une membrane
de nylon (southern blot) et hybridés, soit avec la sonde α marquée, soit
avec la sonde β marquée. Après rinçage, les deux southern sont autoradiographiés
(fig. 11.9).
Vous préciserez la nature et l’amplitude des événements moléculaires
survenus chez certains mutants. Vous préciserez la localisation minimale
du gène STA, à l’échelle moléculaire, sur la carte physique de la région
étudiée.
Solution
1. Le premier croisement sta-1//Y × sta+//sta+ donnent des descendants de génotypes sta+//Y
et sta-1//sta+, viables et fertiles.
Les croisements sta-1//sta-1 × sta-1//Y ou sta-1//sta-1 × sta+//Y ne donnent aucun descendant
car les oeufs ne se développent pas.
Or les zygotes formés par ces deux derniers croisements ont pour génotype sta-1//sta-1
ou sta-1//Y, dans le premier, et sta-1//sta+ ou sta-1//Y, dans le deuxième, dont on sait qu’ils
sont viables puisqu’on en dispose à l’état adulte.
Ces génotypes viables sta-1//sta-1 ou sta-1/Y
n’ont pu être obtenu qu’à partir d’un croisement sta+//sta-1 × sta-1/Y.
On voit qu’on peut toujours obtenir des adultes pour les génotypes sta-1//sta-1, sta-1//Y, ou
sta-1//sta+, à la seule condition que la mère ne soit pas sta-1//sta-1.
Un embryon sta-1//sta-1
peut se développer à partir d’un croisement sta-1//sta+ × sta-1/Y, mais non à partir d’un croisement
sta-1//sta-1 × sta-1//Y.
Que la mutation sta-1 soit une perte de fonction ou qu’elle exerce un effet toxique, on est
amené à conclure que son effet n’est critique pour le développement de l’oeuf qu’en fonction
du génotype maternel et indépendamment du génotype embryonnaire; la mutation sta-1
exerce donc un effet maternel.
Si ce n’était pas le cas, et que le développement de l’oeuf dépendait de l’expression zygotique
du gène STA, le génotype sta-1//sta-1 serait létal, ce qui n’est pas le cas.
On sait qu’un certain nombre d’étapes des premiers stades de développement sont prises en
charge par l’expression de gènes maternels et non de gènes zygotiques.
En effet, l’ovule n’est
pas une cellule isolée, elle est associée à de nombreuses cellules nourricières au sein de
l’ovocyte.
Par transfert de protéines ou de m-RNA, les cellules nourricières de l’ovule apportent
non seulement des réserves nutritives nécessaires aux premiers stades de développement,
mais aussi des produits de gènes nécessaires à ces premiers stades, notamment ceux
qui, du fait de gradients de concentration, génèrent une préformation des axes primitifs de
développement de l’embryon.
On peut donc conclure que le gène STA fait partie de cet ensemble de gènes à effet maternel
dans l’oeuf.
Les oeufs des mères sta-1//sta-1, quel que soit le génotype de l’embryon, sont dépourvus du produit du gène STA qui est censé, non pas être traduit chez l’embryon (sinon
les génotypes sta-1//Y et sta-1//sta-1 seraient létaux), mais chez la mère, durant l’ovogenèse.
Au contraire, le produit du gène STA est présent et actif chez tous les embryons des mères
sta-1//sta+, même les embryons sta-1//sta-1, car il est synthétisé par les cellules nourricières
somatiques sta-1//sta+.
2. Reconstitution d’un protocole permettant l’énoncé des résultats de la question 1.
Le stock est constitué de 1/3 mâles sta-1 viables et fertiles, de 1/3 femelles FM3//sta-1,
viables et fertiles, reconnaissables à leur phénotype [B/2] et de 1/3 femelles sta-1//sta-1,
stériles (ce qu’il convient de démontrer ici), reconnaissables à leur phénotype [B+].
En effet, les croisements fertiles mâles sta-1/Y × femelles FM3//sta-1 donnent 1/4 de zygotes
mâles FM3/Y létaux, 1/4 de zygotes mâles sta-1/Y viables, 1/4 de femelles FM3//sta-1 et
1/4 de femelles sta-1//sta-1.
La fertilité des mâles sta-1/Y est attestée par leur capacité de maintenir la mutation sta-1 en
stock. On peut les croiser avec des femelles sta+//sta+ et constater leur fertilité par l’existence
d’une descendance F1 viable et fertile.
Pour tester la fertilité de femelles sta-1//sta-1, il faut d’abord les produire, ce qui est facile
puisqu’elles sont repérables dans le stock, par le fait qu’elles sont de phénotype [B+].
En les croisant, individuellement dans un tube, avec un mâle sauvage sta+/Y ou un mâle du
stock, sta-1/Y, on peut constater qu’elles pondent des oeufs qui ne se développent jamais.
3. Le croisement d’une femelle FM3//Df par un mâle sta-1//Y permet de récupérer les
femelles F1 de phénotype [B+], c’est-à-dire de génotype sta-1//Df.
L’absence du phénotype
dominant induit par le balanceur étant une indication de la présence de l’autre chromosome,
c’est-à-dire le chromosome porteur de la déficience Df.
Si la délétion du chromosome X couvre le locus du gène STA, les femelles n’ont qu’un seul
exemplaire muté de ce gène et seront stériles; dans le cas contraire, elles seront porteuses
d’un allèle sta+ sur le chromosome Df et seront fertiles.
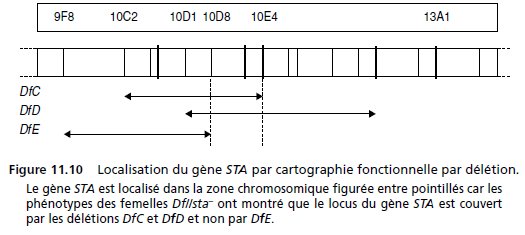
Les résultats montrent que les délétions DfC et DfD couvrent ce locus mais pas la délétion
DfE puisque les femelles sta-1//DfE sont fertiles, ce qui permet de situer le locus du gène
STA entre 10D8 et 10E4 (fig. 11.10)
NB : On devrait dire, en toute rigueur, que le gène est localisé au moins en partie dans cette
zone, car il peut aussi s’étendre à gauche de 10E4.
Cependant, on sait que chez la drosophile il semble y avoir un gène par bande, ce qui permet de dire que le gène STA est bien compris
entre 10D8 et 10E4.
Cette question est une illustration de la méthode de cartographie des gènes par délétion
.
Cependant, au lieu de tester la possibilité ou l’impossibilité d’obtenir des gamètes
sauvages par recombinaison à la méiose, chez l’hétérozygote chromosome muté//chromosome délété, on teste ici directement le phénotype du diploïde.
Remarque.
On ne savait pas a priori si la mutation sta-1 était une « perte de fonction »,
la cartographie fonctionnelle par délétion permet de le démontrer, la déletion des
gènes STA étant une perte de fonction.