4. Aucun des trois marqueurs n’est sur le chromosome X, sinon les deux croisements réciproques
auraient donné des résultats différents, ce qui n’est pas ici le cas.
En effet, dans le cas du premier marqueur m1, si celui-ci était localisé sur le chromosome X,
tous les mâles F1 du premier croisement auraient un profil de type B (chromosome X unique
venant d’une femelle B), alors que les femelles auraient un profil hétérozygote, et dans le
deuxième croisement, tous les mâles F1 auraient un profil de type A (chromosome X unique
venant d’une femelle A), alors que les femelles auraient un profil hétérozygote (fig. 11.4).
5. a L’analyse de la méiose chez les F1 hétérozygotes pour chaque marqueur, montre une
ségrégation 2/2 des « allèles » de ces marqueurs.
Pour le marqueur m3, les effectifs attendus sous l’hypothèse de ségrégation 2/2 sont respectivement
égaux à 100, 100 et 200 (proportions 1/4, 1/4 de chaque homozygote et 1/2 d’hétérozygotes),
ce qui, comparés aux effectifs de 96, 98 et 206, donne un χ2 égal à 0,38,
largement inférieur à 5,99, la valeur qui n’est dépassée par hasard que 5 fois sur cent.
Les
écarts ne sont pas significatifs, on prendrait un risque trop grand de se tromper en rejetant
l’hypothèse de ségrégation 2/2; donc on l’accepte.
5. b Bien évidemment la ségrégation 2/2 est attendue pour un marqueur RFLP, puisqu’on sait
a priori qu’il est, par nature, constitué d’un couple d’« allèles ».
Dans le cas d’un phénotype morphologique ou biochimique, les souches étudiées peuvent
différer, pour ce phénotype, pour un gène ou éventuellement pour plusieurs gènes, et c’est
l’observation d’une ségrégation 2/2 qui nous conduit, mais seulement a posteriori, à la
connaissance du fait que les deux souches étudiées ne diffèrent que pour un seul gène,
mettant en jeu un seul couple d’allèles dans la méiose du F1.
6. Selon la disposition des deux doubles paires de chromatides appariées à la métaphase, on
aura, avec des probabilités égales, soit quatre gamètes parentaux, soit quatre gamètes
recombinés (voir figure 3.1, page 47).
Mais il est aussi possible qu’un crossing-over, survenant entre le locus d’un des deux
marqueurs et son centromère, conduise ce type de méiose à la formation de quatre gamètes,
deux parentaux et deux recombinés.
Mais au bout du compte, sur un grand nombre de méioses, la fréquence des gamètes recombinés
sera égale à celle des gamètes parentaux; l’indépendance physique conduisant à l’indépendance
génétique.
7.a La souche A est :
et la souche B est :
Les pointillés indiquent l’absence de connaissance quant à leur liaison génétique éventuelle.
La souche F1 est donc :
Si les deux marqueurs sont physiquement indépendants, mâles et femelles doivent fournir
quatre types de gamètes équifréquents, notamment des gamètes recombinés (m2+; m3–) ou
(m2–; m3+) qui, selon le hasard des fécondations, pourront conduire à des génotypes F2
doubles homozygotes recombinés, soit (m2+/m2+; m3–/m3–) ou (m2–/m2–; m3+/m3+).
Le premier de ces génotypes présenterait un profil de type A pour le marqueur m2, et de
type B pour le marqueur m3.
Inversement, le deuxième de ces génotypes présenterait un profil de type B pour le
marqueur m2, et de type A pour le marqueur m3.
Or, ce sont deux associations de profil qui ne sont jamais observées. On doit donc en
conclure que ces génotypes n’existent pas et que les gamètes qui pourraient les concevoir
n’ont pas été formés, ce qui est possible chez le mâle, en cas de liaison physique, car il n’y a
pas de crossing-over chez la drosophile mâle.
Bien évidemment des gamètes recombinés (m2+; m3–) ou (m2–; m3+) ont été formés chez
les femelles, sinon les deux marqueurs m2 et m3 auraient coségrégé et on n’aurait eu que
trois classes d’associations de profils, A-A (1/4), B-B (1/4) et F1-F1 (1/2), ce qui n’est pas
le cas.
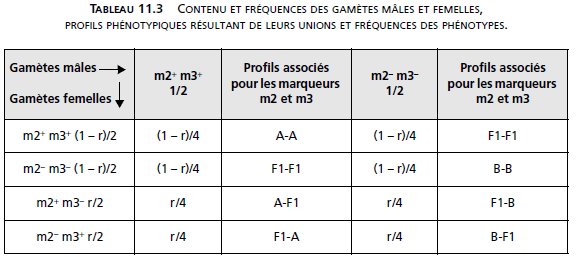
Si on appelle r, la fréquence des gamètes recombinés (par crossing-over) chez les femelles, il
est alors possible de calculer la fréquence théorique des différents phénotypes du tableau,
après avoir calculé la fréquence théorique de chacun des génotypes résultant des croisements
F1 × F1, en fonction de r/2, la fréquence de chacun des types de gamètes recombinés,
et (1 – r)/2, la fréquence de chacun des gamètes parentaux (tabl. 11.3).
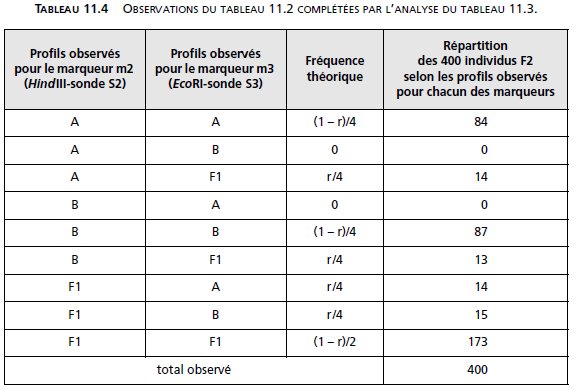
D’où le tableau d’analyse tenant compte des fréquences théoriques des phénotypes en fonction
de r.
ce qui permet d’estimer facilement r : r = (14 + 13 + 14 + 15)/400 = 56/400 = 0,14.
Les marqueurs m2 et m3 sont physiquement et génétiquement liés, à une distance égale à
14 unités de recombinaison.
7. b Dans le cas où les marqueurs auraient été génétiquement indépendants, deux situations
cartographiques différentes auraient été possibles, les deux marqueurs pouvaient être physiquement
indépendants ou physiquement liés.
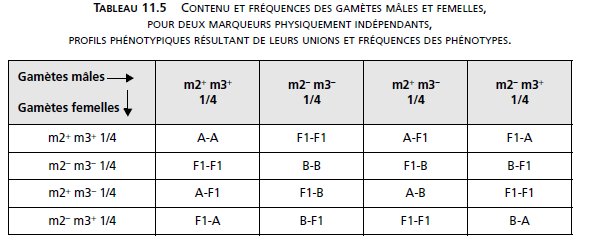
Dans le cas d’indépendance physique, le mâle fournit quatre types de gamètes équifréquents,
comme la femelle (tabl. 11.5).
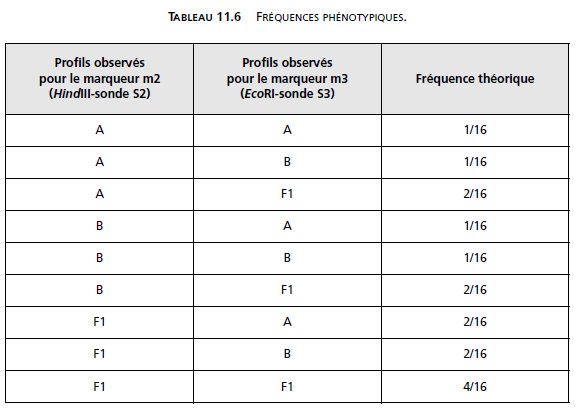
Ce qui conduirait à l’observation des fréquences phénotypiques suivantes (tabl. 11.6).
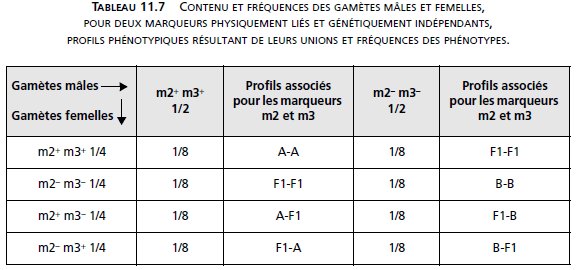
Dans le cas d’indépendance génétique avec liaison physique, les femelles donnent quatre
types équifréquents de gamètes, mais les mâles ne fournissent que des gamètes parentaux
(tabl. 11.7).
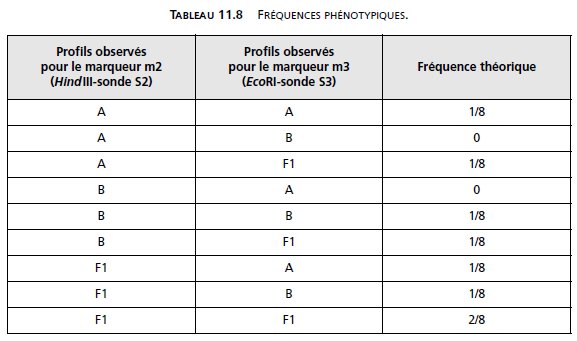
Ce qui conduirait à l’observation des fréquences phénotypiques suivantes (tabl. 11.8).
NB : Le tableau 11.8 est l’équivalent du tableau 11.4, avec r = 1/2.