1.a. Le diploïde formé est muté ponctuellement dans le gène E du chromosome et délété pour
le gène A (éventuellement B, sans toucher le site m5) sur l’épisome.
Ce diploïde est donc pourvu de toutes les séquences codantes de l’opéron mais il n’y aura
complémentation fonctionnelle que si tous ces gènes sont exprimés.
Ce ne serait pas le cas
dans deux situations :
– soit parce que la délétion toucherait le promoteur, si celui-ci est situé du côté de TYR, ce
qui bloquerait l’expression de E sur l’épisome; E étant muté sur le chromosome, on serait
en situation de non complémentation;
– soit parce que E serait une mutation polaire et bloquerait alors l’expression des gènes en
aval, si le promoteur est du côté de PUR, notamment le gène A qui est délété sur l’épisome,
ce qui conduirait à une situation de non complémentation pour ce gène.
Le phénotype observé [trp+] n’étant possible qu’en raison de la complémentation fonctionnelle,
on peut en déduire que les gènes E de l’épisome, et A du chromosome, sont actifs et qu’en conséquence, le promoteur est obligatoirement du côté de PUR et que la mutation m3
n’est pas polaire.
b. On étale les conjugants sur un milieu Mo additionné de streptomycine pour contre-sélectionner
les donatrices, avec tyr mais sans pur, pour sélectionner les réceptrices de l’épisome
qui leur confère le phénotype [pur+; tyr–].
On ajoute également du tryptophane puisqu’il
n’est pas certain qu’il y ait complémentation fonctionnelle, et on teste le phénotype trp par
réplique sur un milieu dépourvu de trp; donc milieu Mo + tyr + trp + str.
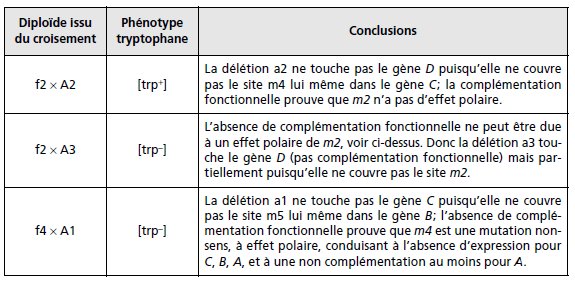
2. Réponse troisième colonne du tableau.
3.a. L’activité spécifique du complexe D/E passe de 100 à 20 quand on ajoute du trp dans le
milieu réactif, ce qui prouve que le trp est un inhibiteur de l’activité enzymatique D/E.
En présence de trp, l’ensemble de la chaîne de biosynthèse est ralenti par inhibition rétroactive
du produit terminal, ici le trp, sur le premier « maillon » de cette chaîne.
Le tryptophane joue aussi le rôle d’inhibiteur dans la régulation transcriptionnelle de
l’opéron puisqu’en culture en présence de trp, les quantités de protéines produites sont divisées
par 50 (on passe de 100 à 2).
b. Soit le tryptophane joue son rôle d’inhibiteur transcriptionnel en se fixant à un activateur
de l’opéron pour le rendre inactif; soit le tryptophane joue son rôle d’inhibiteur transcriptionnel
en se fixant à un répresseur de l’opéron pour le rendre actif.
4.a. Le mutant constitutif peut être muté dans le gène de structure de l’activateur ou du
répresseur, mais peut aussi être un mutant de cible sur l’opéron trp.
Si c’est un mutant dans le gène régulateur et que ce gène est localisé dans une autre partie du
génome que l’opéron trp, le croisement entre une Hfr sauvage et la souche constitutive réceptrice
pourrait permettre à celle ci de retrouver sa capacité de régulation, c’est-à-dire de perdre
sa constitutivité, par remplacement de son gène muté par la séquence sauvage exogène
apportée par la Hfr.
Les trois Hfr utilisées permettent en 50 minutes de parcourir une moitié ou une autre du
génome et de tester la présence du gène régulateur sauvage par la capacité de retrouver des
non constitutifs parmi les réceptrices recombinantes pour un marqueur distal de recombinaison.
Le croisement avec la Hfr1 montre que la constitutivité demeure et que le gène régulateur
n’est pas situé entre l’origine et le marqueur his.
La capacité de perdre la constitutivité dans les deux autres croisements montre que le gène
régulateur est dans la partie commune des gènes transférés par Hfr2 et Hfr3, à l’exclusion de
ceux transmis par Hfr1, soit la zone génomique comprise entre 75 et 100 mn, et sans doute
très près du marqueur sérine puisque seuls 5 % des recombinants ont acquis la séquence ser+
sans co-acquérir le gène régulateur sauvage.
La souche Bc est constitutive par mutation dans un gène régulateur de l’opéron tryptophane,
mais la localisation d’un tel gène ne peut permettre de statuer sur sa fonction; il peut spécifier
un activateur ou un répresseur…
b. Le plasmide permet de constituer un diploïde partiel pour le gène régulateur, sélectionnable
sur Mo en présence de kanamycine, additionné des trois acides aminés correspondant
aux phénotypes d’auxotrophie et de tryptophane pour tester le maintien de la constitutivité
ou sa perte par le retour à la régulation qui conduiraient respectivement au maintien d’une
activité spécifique élevée pour la typtophane synthétase ou à une chute de celle-ci.
L’observation d’une telle chute prouve que la séquence mutée a un effet récessif par rapport
à la séquence sauvage, ce qui plaide en faveur d’une régulation négative de l’opéron trp.
Dans un tel cas, la mutation constitutive peut être une perte de fonction dans le gène d’un
répresseur, compensable par l’allèle sauvage chez le diploïde.
Si le gène régulateur spécifiait un activateur, la constitutivité serait due, non pas à une perte
de fonction mais à une perte d’affinité pour le trp, bloquant ainsi son action inhibitrice; on
s’attendrait alors à un effet dominant de l’allèle muté sur son homologue sauvage (ou au
moins partiellement dominant), ce qui ne correspond pas aux observations.
5.a. La zone 1 est le site de fixation du répresseur. L’effet phénotypique d’une délétion est
donc identique à celui d’une perte de fonction dans le gène de ce répresseur : l’opéron est
transcrit de manière constitutive par absence d’action du répresseur que celui-ci soit absent
ou qu’il ne puisse de fixer et agir.
L’apport du répresseur par le plasmide ne peut avoir aucun
effet en absence de son site de fixation.
b. La souche Bc est constitutive, elle transcrit et traduit son opéron en absence ou en
présence de trp qui ne peut plus exercer son rôle d’inhibiteur transcriptionnel via sa liaison
au répresseur.
L’absence de ce rôle est attestée par l’égale efficacité d’initiation de la transcription puisqu’en
présence ou en absence de trp, les quantités de messagers initiés sont égales.
Cependant en absence de trp tous les messagers initiés sont complètement terminés et
traduits alors qu’en présence de trp 100 % des transcriptions progressent jusqu’à la zone 3 et
seules 10 % d’entre elles continuent et permettent d’exprimer la totalité de l’opéron.
La présence ou l’absence de tryptophane affecte la transcription de l’opéron après que le
messager ait commencé à être traduit puisque dans tous les cas on a une quantité égale de
messager transcrit et traduit pour le peptide leader.
Le phénomène mis en évidence concerne la progression de la transcription et non son initiation
et peut donc être expliqué par la présence d’une séquence terminatrice de la transcription
active uniquement en présence de trp.
Et la délétion de cette séquence terminatrice rétablit la capacité de progression de la transcription
en présence de trp.
Pour pouvoir mettre en évidence l’effet du trp sur la progression de la transcription, il est
nécessaire de s’affranchir de son effet sur l’initiation, ce qui justifie l’étude chez un mutant
constitutif par absence de répresseur.
c. On pourrait formellement imaginer que la séquence de la zone 3 soit le site de fixation
d’un activateur ou d’un autre répresseur.
Dans ce cas on devrait pouvoir obtenir des mutants
du gène de structure de cet activateur hypothétique, ce qui n’est pas le cas.
Par ailleurs les
observations antérieures montrent que le phénomène observé affecte la progression de la
transcription et non son initiation, ce qui n’est pas non plus compatible avec l’action d’un
activateur qui agit sur l’initiation de la transcription.
Ce phénomène qui affecte non l’initiation de la transcription, mais sa progression et son arrêt
(dans une majorité de cas) en raison de l’interaction avec le messager, en présence de tryptophane
a reçu le nom d’atténuation.