4.a. Ce croisement permet le passage de l’épisome dans les F–; le milieu d’étalement permet
de récupérer des colonies de phénotype [pur+, tyr+] qui ne peuvent être de doubles révertants
mais des réceptrices ayant acquis un épisome qui leur confère ce phénotype, ce qui atteste
que les allèles mutés d’auxotrophie pour les purines et la tyrosine ont un effet récessif
compensable par celui des allèles sauvages.
La réplique permet de tester la dominance ou la récessivité de la mutation de résistance au
phage; celle ci est récessive par rapport à l’allèle de sensibilité.
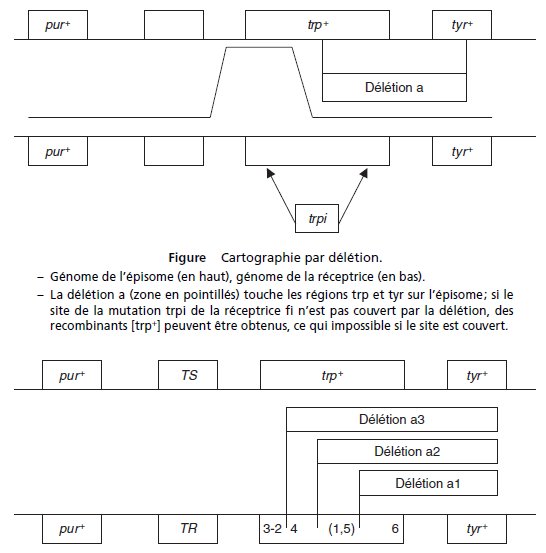
b. On sait que les rayons X induisent des délétions et on récupère des souches mutées à la fois
dans un (ou plusieurs) gène(s) gouvernant la synthèse du tryptophane et un gène gouvernant
celle de la tyrosine, dont on sait par ailleurs qu’ils sont contigus; il s’agit donc très vraisemblablement
de mutants par délétion affectant une partie de l’épisome dans les régions trp et tyr.
Les croisements réalisés permettent d’effectuer une cartographie par délétion des sites de
mutation, sachant qu’il est possible (voir figure) de reconstituer un gène sauvage si et seulement
si le site de mutation ponctuel (dans le génome f1 à f6) n’est pas couvert par la délétion
(a1 à a3) apportée par un autre génome ou fragment de génome (épisome).
À partir de ce schéma appliqué aux résultats des croisements, on peut cartographier les sites
et l’ampleur des délétions :
L’ordre des mutations 1, 2 et 3 a été déterminé par conjugaison et est ici confirmé, les mutations
4 et 6 sont cartographiées mais l’ordre des mutations 1 et 5 reste indéterminé.
c. Les mutants obtenus sont des mutants ponctuels dans la région trp de l’épisome et le croisement
avec les souches f1 à f6 permet de récupérer des réceptrices [tyr+] ayant acquis l’épisome
et de tester la complémentation fonctionnelle après s’être assuré, par la formation du diploïde
(F + épisome muté) que les mutations trp– de l’épisome avaient un effet récessif compensable
par celui de l’allèle sauvage du gènome de F.
On peut alors définir cinq groupes de complémentation, ce qui permet de considérer que
l’opéron tryptophane contient au moins cinq gènes, (Ae, 3), (Ad, 2), (Ac, 1,4), (Ab, 5) et (Aa, 6).
les mutations 1 et 4 touchant le même gène et sachant que les mutations 1 et 5 sont couvertes par
une même délétion, on peut en déduire l’ordre : trp4-trp1-trp5, et l’ordre final : 3-2-4-1-5-6.
5. L’utilisation du phage dans le milieu d’étalement permet d’être assuré que les colonies
obtenues ont bien recombiné, en délaissant le gène TS de l’épisome et en reconstruisant une
région trp sauvage, démontrant alors que la délétion ne couvre pas le site ponctuel.
En effet, en l’absence de phages, des diploïdes pourraient éventuellement pousser en cas de
complémentation fonctionnelle, si la délétion ne touche pas le ou les même(s) gène(s) que la
mutation trpi, ou bien d’éventuels révertants pour la mutation ponctuelle trp.
Problème 12.9
Étude génétique de l’opéron tryptophane (deuxième partie)
À la suite des mutagenèses et des diverses expérimentations de recombinaison
réalisées dans le début de l’étude (exercice précédent), on dispose
de six souches F, notées f1, f2, f3, f4, f5 et f6 de phénotype [pur–; TR; trp–; tyr–; strpR] porteuses d’une mutation ponctuelle dans un gène de l’opéron,
dont la position et l’ordre ont été définis précédemment.
On rappelle que
ces six mutations ont un effet récessif par rapport à leurs homologues
sauvages respectifs.
On dispose également d’une souche A [strpS], contenant un épisome
porteur de la séquence sauvage des gènes PUR, T, TRP et TYR, à partir de
laquelle on a obtenu des mutants A1, A2 et A3, également caractérisés par
rapport aux mutants f et à l’opéron tryptophane.
Question 1.
Par un croisement approprié entre f3 et A1, on récupère un diploïde partiel
pour l’opéron tryptophane.
a. Précisez quel est le phénotype tryptophane attendu en fonction de la
position du promoteur et/ou de la nature de la mutation m3.
Le phénotype
observé étant [trp+], que pouvez vous en conclure ?
b. Sur quel milieu est-il préférable de récupérer les diploïdes après la
conjugaison ? justifiez votre réponse.
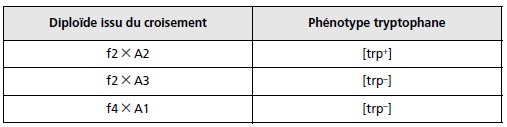
Question 2.
On construit d’autres diploïdes dont on teste le phénotype tryptophane
(tableau); interprétez les résultats obtenus en précisant, quand cela est
nécessaire et possible, l’amplitude des délétions et la nature des mutations
ponctuelles ainsi testées.
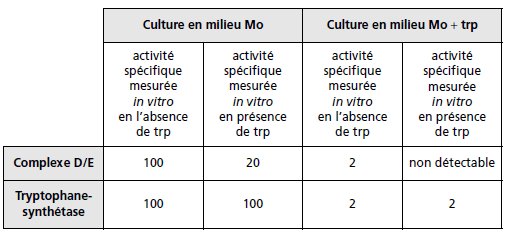
Question 3.
Les chaînes peptidiques codées par les gènes D et E s’unissent pour former
un complexe enzymatique, noté D/E, assurant la première étape de la chaîne
de biosynthèse du tryptophane, la chaîne peptidique codée par le gène C
assure la deuxième étape tandis que les chaînes peptidiques codées par les
gènes A et B s’unissent pour former la tryptophane-synthétase, un hétérotétramère
(deux chaînes A et deux chaînes B) assurant la troisième et dernière
étape.
On réalise deux cultures d’une souche sauvage en milieu Mo ou Mo + trp,
puis après extraction des protéines on réalise un dosage spécifique de D/E
et de tryptophane-synthétase, en présence ou en absence de trp.
Le tableau suivant rapporte les valeurs de l’activité spécifique mesurée dans
chacun des cas.
a. En justifiant vos conclusions, vous montrerez que le tryptophane agit à
deux niveaux pour réguler sa propre synthèse.
b. Vous proposerez deux schémas différents de la régulation transcriptionnelle
de l’opéron tryptophane.
Question 4.
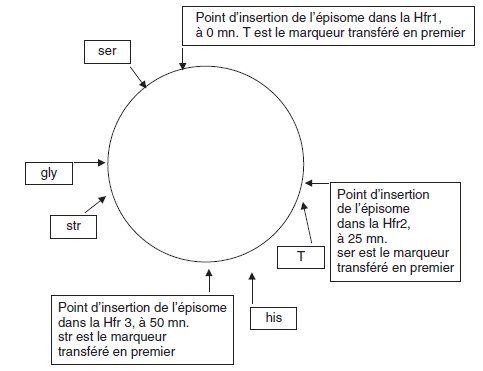
À partir d’une souche B [ser–, TR, trp+, his–, gly–, strR] on a obtenu un
mutant constitutif, noté Bc, capable de produire des quantités importantes de complexe D/E ou de tryptophane-synthétase même en culture sur Mo +
trp (les locus des mutations ponctuelles d’auxotrophies sont placées sur la
carte jointe).
a. On réalise, pendant 50 minutes, trois conjugaisons parallèles entre la
souche Bc et chacune des trois Hfr de génotype sauvage, Hfr1, Hfr2 ou Hfr3
(voir sur la carte, la localisation et le sens de l’origine de transfert).
1 %0% des colonies [his+, strR] issues du croisement Bc × Hfr1 sont constitutives
et donnent une activité spécifique élevée de tryptophane synthétase
après culture en milieu Mo + trp.
%0% des colonies [gly+, strR] issues du croisement Bc × Hfr2 sont constitutives
et 5 % des colonies [ser+, TR] issues du croisement Bc × Hfr3 sont
constitutives.
Interprétez ces résultats sur le plan cartographique et fonctionnel en précisant
si cela vous permet de choisir entre les deux modèles de régulation de
l’opéron tryptophane.
b. La séquence sauvage de la région porteuse du gène, précédemment
identifié, a été clonée dans un plasmide conférant la résistance à la kanamycine.
On transforme la souche Bc par ce plasmide; les transformées [kanR] cultivées
sur Mo + sérine + tryptophane + glycine + histidine fournissent des quantités
à peine dosables de tryptophane-synthétase après extraction protéique.
En faveur de quel modèle de régulation ce résultat plaide-t-il ? justifiez
votre réponse.
Question 5.
Une étude moléculaire fine du promoteur de l’opéron tryptophane a été
réalisée et permet d’y définir plusieurs séquences ou zones de fonction
différentes :
a. On a montré que la délétion de la zone 1 (voir schéma) conduisait à un
phénotype identique à celui de Bc.
La transformation avec le plasmide défini à la question précédente ne
modifie pas le phénotype constitutif. Concluez.
b. On a découvert qu’outre les cinq chaînes peptidiques des gènes de structure,
l’opéron codait aussi pour une chaîne peptidique « leader » de 14 acides
aminés sans fonction enzymatique (la zone 3 est assez courte pour que l’arrêt de la traduction au codon STOP de la séquence leader ne bloque pas la
transcription en aval).
En voulant étudier finement la régulation transcriptionnelle par la mesure
de la longueur des messagers, on a fait les observations suivantes :
– le mutant Bc cultivé en milieu Mo + ser + his + gly produit une quantité de
messager constituée presque exclusivement de messagers de 7 000 nucléotides
correspondant à l’expression de tous les gènes de l’opéron;
– le mutant Bc cultivé sur même milieu additionné de trp, produit une
quantité égale de messagers mais constitués pour 10 % de messagers de
7 000 et pour 90 % de messagers de 130 nucléotides correspondant
environ aux zones 1 & 2;
– la délétion de la zone 3 redonne au mutant Bc la capacité de produire, en
milieu Mo + trp, 100 % de messagers longs.
Montrez que ces résultats sont compatibles avec l’hypothèse d’une séquence
terminatrice de la transcription localisée dans la zone 3 et active en présence
de tryptophane.
La solution vous étant donnée, il vous est simplement demandé de présenter
une argumentation convenablement construite, justifiant notamment le génotype
de la souche utilisée dans cette expérimentation.
c. Quelle autre hypothèse aurait pu, au moins formellement, expliquer la
conséquence de la délétion de la zone 3 ?
En quoi le fait qu’aucune autre mutation du génome en dehors de la délétion
de la zone 3 ne puisse avoir le même effet que cette délétion prouve
que cette hypothèse ne peut pas être conservée ?
En quoi les deux autres observations ne sont-elles pas non plus très compatibles
avec cette hypothèse ?