4. Le fragment cloné dans pCOS2 a une taille comprise entre 33 et 49 Kb, il ne peut donc pas
être cloné dans un phage λ.
Il est découpé et sous-cloné afin d’isoler le gène sodA dans un
petit fragment cos2, susceptible d’être cloné dans λts.
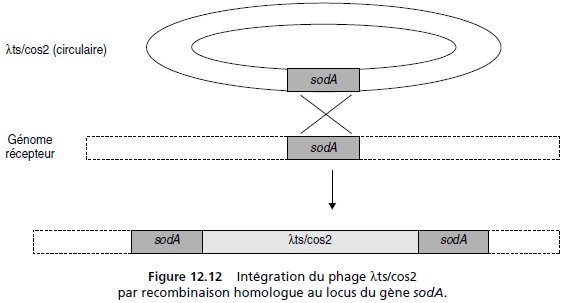
Le clonage de sodA, à la place des séquences d’attachement et de l’intégrase, fait perdre au
phage sa capacité de lysogénie, à moins que le phage ne soit intégré au génome bactérien par
recombinaison homologue au locus du gène sodA (fig. 12.12), ce qui est le but recherché.
En effet, les plages troubles contiennent des bactéries lysogènes, dont on montre, après les
avoir purifiées par réisolement, qu’elles présentent l’induction lytique à 42 °C, caractéristique
du phage mutant λts utilisé.
Ce phage est donc intégré au locus du gène sodA.
Il est désormais possible de cartographier le gène sodA en cartographiant le locus d’insertion
du phage λts/cos2 dans la souche B1, c’est-à-dire en cartographiant la thermosensibilité.
L’intégration du phage λts/cos2 au locus du gène sodA conduit à la duplication de ce gène
(fig. 12.12) ce qui explique le doublement de l’activité dans le dosage, alors que la souche A2
contenait en moyenne trois copies du cosmide, donnant une activité quatre fois supérieure.
5. On sélectionne respectivement, à 40 minutes, dans chacun des croisements, les recombinants
[trp+], [pro+], [pro+] et [ilv+]. Chez chacun de ces recombinants, la séquence résidente
trp–, ou pro–, ou ilv– a été remplacée par la séquence sauvage trp+, ou pro+, ou ilv+, transférée
par la Hfr.
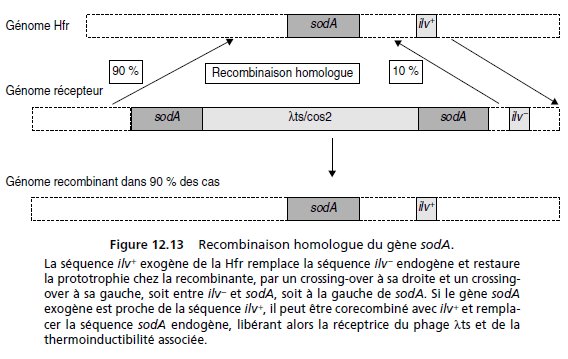
Par ailleurs, le gène sodA de la Hfr peut être transféré à la réceptrice et y remplacer la copie
endogène.
Cela ne change rien pour l’information génétique contenue dans les copies
exogène (Hfr) et endogène (réceptrice) de sodA qui sont toutes deux sauvages, mais, dans
cette recombinaison, le gène sodA endogène peut quitter le génome récepteur avec le
phage λts qui lui est adjacent et qui ne présente pas de séquence homologue sur le fragment
de génome venant de la Hfr (fig. 12.13).
Dans ce cas le recombinant, au locus du gène sodA, ne sera plus lysogène, ce qui se traduira
par la perte de l’inductibilité lytique à 42 °C et la capacité de croissance à cette température.
Si, de plus, le gène sodA est proche d’un des locus testés pour la prototrophie (pro, ilv
ou trp), la séquence du gène sodA peut être cotransférée et corecombinée; dans ce cas, le
recombinant sauvage prototrophe aura de fortes chances d’être, en même temps, devenu
thermorésistant (fig. 12.13).
À l’inverse, si le gène sodA est loin de la séquence recombinante testée (pro, ilv ou trp), le
phage λts/cos2 a très bien pu, même si le gène sodA a été transféré, ne pas avoir été concerné
par la recombinaison moléculaire.
Bien évidemment, si le gène sodA n’est jamais transféré,
tous les recombinants prototrophes testés auront gardé le phage λts/cos2 et seront restés
thermoinductibles et ne pourront croître à 42 °C.
Ainsi, en testant la perte de l’inductibilité lytique (thermosensibilité) chez des recombinants
prototrophes, en fonction de l’origine de transfert de la Hfr, on peut localiser le phage λts/cos2
et donc le gène sodA.
Les résultats avec les Hfr1 ou 3 montrent que le gène sodA n’est jamais cotransmis (100 %
des recombinants restent thermoinductibles et ne peuvent croître à 42 °C), ce qui implique que le gène sodA ne réside pas entre les sites d’insertion de l’épisome de ces deux Hfr (fragment
contenant leu, pro, trp).
Les résultats avec les Hfr2 ou 4 montrent que le gène sodA est transmis (des recombinants ne
sont plus thermoinductibles et peuvent croître à 42 °C), ce qui implique que le gène sodA
réside entre le site d’insertion de l’épisome de la Hfr2 et celui de la Hfr1 (fragment contenant
ilv).
Le pourcentage supérieur de recombinants thermorésistants avec la Hfr4, où on teste les
recombinants [ilv+], implique que le gène sodA est très proche de ilv et qu’il est corecombiné
dans 90 % des cas (voir fig. 12.13 les deux modalités de recombinaison moléculaire conduisant
soit à 10 % de thermoinductibles soit à 90 % de thermorésistant parmi les [ilv+]).
Avec la Hfr2, le gène sodA est toujours transféré, mais on teste les recombinants [pro+] qui
peuvent très bien ne pas avoir recombiné pour la séquence porteuse du gène sodA, ce qui
explique la diminution de la fréquence de thermorésistants parmi les [pro+].
6. Ce résultat implique que le gène sodB est compris entre les sites d’insertion de l’épisome
des Hfr2 et 3 (fragment contenant le locus str).
On comprend, dès lors, que la carte de restriction
de la séquence pCOS3 n’ait aucun rapport avec celle du fragment de l’épisome F′1.
Problème 12.8
Étude génétique de l’opéron tryptophane (première partie)
On dispose :
– d’une souche HfrH de coli, porteuse, à 23 mn de son origine de transfert,
d’une mutation, notée pur–, responsable d’un phénotype d’auxotrophie
pour les purines et d’une mutation, notée TR, conférant une résistance à
un phage T,
– d’une souche F– porteuse d’une mutation, notée tyr–, responsable d’un
phénotype d’auxotrophie pour la tyrosine, et d’une mutation, notée strR,
conférant la résistance à la streptomycine.
Question 1.
Les souches H et F sont croisées et on réalise un étalement toutes les
minutes de deux échantillons, après séparation des conjugants, le premier
sur une boîte de milieu Mo + pur + tyr + phage T + str, le deuxième sur une
boîte de milieu Mo + pur + str.
a. des colonies apparaissent sur le premier milieu à la 25e minute et sur le
deuxième milieu à la 27e, interprétez et concluez.
b. 40 % des colonies obtenues sur le premier milieu d’étalement sont capables
de pousser sur Mo + tyr + T + str, et 60 % des colonies obtenues sur le
deuxième milieu sont capables de pousser sur Mo + pur + T + str, interprétez
et concluez.
Question 2.
On obtient, à partir de la souche F, six mutants indépendants, auxotrophes
pour le tryptophane et notés f1 à f6.
Toutes ces souches, croisées avec la Hfr, donnent des colonies sur un
milieu Mo + pur + tyr + str à partir de la 25e minute.
On transduit alors la souche H par un lysat transducteur préparé sur la souche
f1 et on recueille les recombinants [pur+]; ceux-ci sont testés par réplique,
ce qui permet de dénombrer 410 [TR, trp+], 120 [TS, trp–], 50 [TS, trp+]
et 5 [TR, trp–].
Des résultats semblables sont obtenus à partir des lysats des autres souches
f2 à f6.
Interprétez ces expérimentations et concluez.
Question 3.
On récupère à partir des boîtes d’étalement des transductants (question
précédente) les Hfr [TR, trp–]; elles sont respectivement notées h1 à h6
selon qu’elles sont issues d’un lysat transducteur préparé sur f1, f2, f3, f4,
f5 ou f6.
a. Quel est leur phénotype pour la tyrosine ? Justifiez votre réponse.
b. On croise la souche h2 par f1 pendant 30 minutes et on étale sur un
milieu Mo + tyr + str; parmi les colonies obtenues, 8 % sont capables de
pousser après avoir été répliquées sur un milieu identique additionné de
phages T.
Interprétez ce résultat et montrez sa cohérence quantitative avec celui de la
question précédente.
Justifiez l’apport de tyrosine.
c. On croise la souche h2 par f3 pendant 30 minutes et on étale sur un
milieu Mo + tyr + str; parmi les colonies obtenues, 92 % sont capables de
pousser après avoir été répliquées sur un milieu identique additionné de
phages T.
Interprétez ce résultat.
Question 4.
On dispose d’une souche A, délétée tous les gènes allant de pur compris à
tyr compris, mais porteuse d’un épisome F′ contenant la séquence sauvage
de ces mêmes gènes.
a. On isole, à partir du premier milieu d’étalement des conjugants H × F
(voir question 1), une colonie de phénotype [pur–, tyr–]; cette colonie est
croisée avec A; les étalements sur un milieu Mo + str donnent des colonies
incapables de pousser après réplique sur un milieu identique additionné de
phage T; concluez.
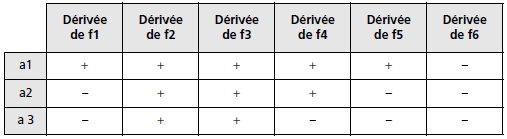
b. On traite la souche A aux rayons X et on étale sur un milieu Mo + tyr
+ trp; parmi les colonies qui poussent, on isole par réplique, trois mutants
de phénotype [tyr–, trp–, TS], notés a1, a2 et a3.
On croise chacun de ces trois mutants avec des dérivées [tyr+, pur+, trp–,
TR] de f1 à f6, obtenues par conjugaison avec une HfrH.
On effectue les étalements sur un milieu Mo + T + str; le tableau suivant
indique les croisements à l’issue desquels on peut observer des colonies
(signe +) et ceux à l’issue desquels on n’en observe pas (signe –).
Interprétez
ce résultat.
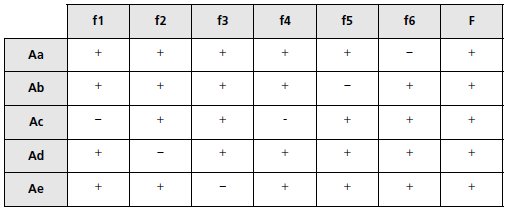
c. On cultive la souche A en présence d’un agent mutagène favorisant les
substitutions de bases et on isole cinq mutants [trp–], notés Aa, Ab, Ac, Ad
et Ae. On croise ces mutants avec les souches f1 à f6 et F; on étale sur milieu
Mo + trp + str, puis on réplique sur Mo + str ; le tableau suivant indique
la présence (+) ou l’absence (-) de colonies sur les boîtes de réplique;
concluez.
Ces résultats doivent vous permettre de compléter la cartographie des
mutations, d’établir celle des gènes de l’opéron tryptophane. Justifiez vos
réponses.
Question 5.
Justifiez l’utilisation du phage dans le milieu d’étalement, dans la question
précédente 4-b afin de pouvoir procéder à l’interprétation des résultats.
Solution
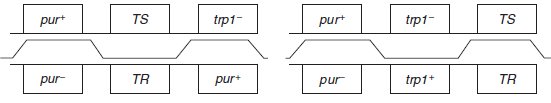
1.a. Les gènes sont dans l’ordre pur (23 mn)- TR (25 mn)-tyr (27 mn).
b. Les 40 % de colonies poussant sur la réplique sont [pur+, TR], ce qui indique qu’il y a eu
60 % de co-recombinaison [pur–, TR], ce qui, compte tenu des distances (2 mn entre pur et TR,
et 2 mn entre TR et tyr) est équivalent à la corecombinaison [tyr+, TR].
2. les mutations sont toutes à 25 mn de l’origine de transfert, c’est-à-dire entre pur et tyr, au
voisinage du site de mutation de la résistance au phage, avec deux ordres possibles (figure).
Les recombinants minoritaires attendus dans chaque ordre, exigeant quatre recombinaisons,
sont respectivement [pur+, TR, trp–] et [pur+, TS, trp+].
L’observation du recombinant minoritaire [pur+, TR, trp–] permet de valider l’ordre 1 et de
rejeter l’ordre 2.
Les recombinants entre les sites pur et T sont les plus nombreux, ce qui indique que le
domaine 2 est le plus grand, suivi du domaine 4, puis du domaine 3 dans lequel on recombine
55 fois sur 645 (environ 8 %).
Les cinq autres mutations, chez f2 à f5, sont situées du même côté du site TS.
3.a. Elles sont [tyr+] car la mutation tyr– ne peut être cotransduite avec la séquence pur+ qui est
à 4 mn, soit à 160 000 pb, alors que le phage transducteur ne peut encapsider que 100 000 pb.
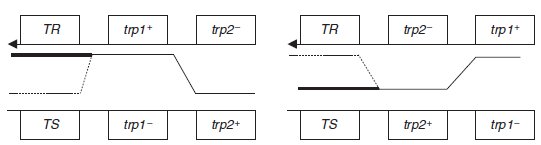
b. La streptomycine contre-sélectionne les donatrices Hfr ainsi que l’absence de purine et
sélectionne les réceptrices recombinantes [pur+, trp+], la réplique sur un milieu additionné de
phages permet de tester l’importance de la co-recombinaison de TR, importante si le site muté de la Hfr est distal par rapport au site muté de la réceptrice (figure de gauche), et très
rare s’il est proximal (figure de droite).
Le croisement peut être figuré ainsi en fonction des deux ordres possibles des sites de mutation trp :
8 % des recombinants [trp+] sont [TR] ce qui indique que 92 % sont [TS], ce qui valide
l’ordre 2 où le site muté de la Hfr est proximal par rapport à celui de la F.
Ce résultat est quantitativement cohérent avec celui de la question précédente où on peut
estimer qu’il y a 8 % de recombinants entre les sites T et trp1.
On ajoute de la tyrosine au milieu afin de s’affranchir de la recombinaison ou non de tyr+; ici
les sites trp jouent à la fois le rôle de marqueur distal de sélection et de marqueurs étudiés.
c. Ce croisement permet de montrer que le site muté sur la Hfr est distal par rapport au site
muté sur la réceptrice, ce qui donne l’ordre suivant pour les trois sites : trp3-trp2-trp1.