Problème 12.7
On connaît chez coli deux activités superoxydes dismutases (sodA et sodB)
dont la fonction est de réduire les radicaux peroxydiques très pathogènes
pour l’intégrité cellulaire, voire létaux.
L’activité sodA fonctionne en
présence de Mn++ et l’activité sodB, en présence de Fe++.
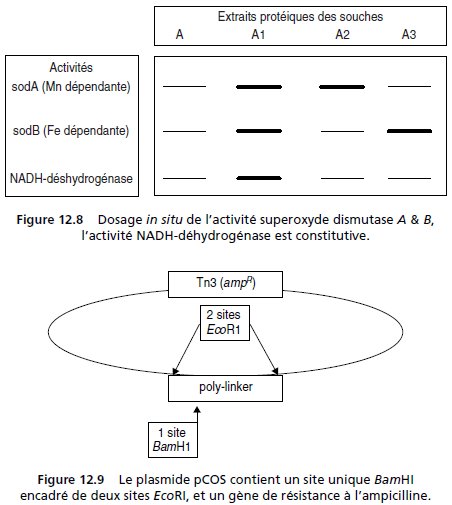
Le dosage simultané des deux activités peut être réalisé in situ sur un gel
d’agarose, après électrophorèse d’un extrait protéique acellulaire (fig. 12.8,
première piste).
1. Il a été impossible d’obtenir des mutants déficients dans l’une ou l’autre
de ces deux activités.
Avez-vous une explication simple ? Dites pourquoi il est, dès lors, impossible
de localiser les gènes codant ces deux activités.
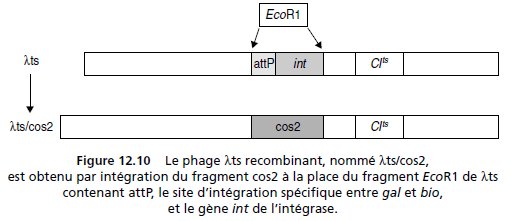
2. On digère partiellement l’ADN d’une souche sauvage de coli par
l’enzyme BamH1 et on récupère, après électroélution sur un gel d’électrophorèse,
les fragments compris entre 33 et 49 Kb, afin de construire des
cosmides recombinants avec le cosmide pCOS (fig. 12.9).
Ces cosmides,
empaquetés in vitro, permettent de transformer une souche réceptrice A de
phénotype [F–; leu–; strR, recA–], étalée sur milieu complet avec ampicilline.
NB : Un cosmide est un plasmide porteur des séquences terminales
« COS » du génome du phage λ.
On peut y intégrer des fragments d’ADN
assez longs et procéder à un encapsidage in vitro permettant alors la pénétration
du cosmide dans une bactérie via la capside.
Une fois entré le
cosmide se recircularise grâce aux séquences COS et se comporte comme
un plasmide, avec, en général plusieurs copies par cellules.
Les colonies isolées sont reprises une à une et mises en minicultures; puis,
après lyse et extraction protéique, on procède à une électrophorèse suivie
d’une révélation in situ de l’activité superoxyde dismutase, dont les résultats
sont rapportés pour trois transformants ampR, nommés A1, A2 et A3
(fig. 12.8, pistes 2, 3 et 4).
Par ailleurs, le transformant A3 présente aussi
un phénotype [leu+].
a. Quel est le but de ce protocole ?
Justifiez son choix, ainsi que le phénotype recA– de la réceptrice.
b. Interprétez les observations électrophorétiques.
Le transformant A1 estil
intéressant ?
3. La carte de restriction du fragment cloné dans pCOS3 (cosmide de A3)
ne se superpose à aucune sous-partie de la carte de restriction de la région
sauvage contenue dans l’épisome F′1 (fig. 12.11).
De plus, la transduction d’une souche A′ de phénotype [F–; leu–; strR,
recA+], par un lysat de P1 transducteur, préparé à partir de A3, se révèle
incapable de produire des recombinants [leu+].
Que peut-on en conclure, sur le plan cartographique et fonctionnel, pour la
région clonée dans pCOS3, compte tenu du phénotype du transformant A3 ?
Justifiez vos réponses, en expliquant clairement l’échec de la transduction.
4. Le fragment cloné dans le plasmide pCOS2 (cosmide de A2) est
découpé par EcoR1 en petits sous-fragments.
L’un d’eux, noté cos2, cloné
dans un plasmide multicopies, confère un accroissement considérable de
l’activité sodA aux bactéries transformées par ce plasmide.
a. On établit alors le protocole suivant :
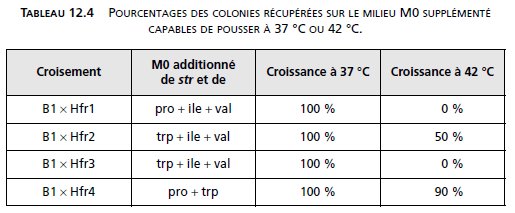
– Le petit fragment cos2 est isolé et cloné dans un phage λts (fig. 12.10)
pour obtenir le phage recombinant nommé λts/cos2.
NB : Le phage λts est porteur d’une mutation CIts, mutation thermosensible
du gène CI entraînant l’induction lytique chez les bactéries lysogènes,
dès qu’elles sont cultivées à 42 °C.
CI, le répresseur de λ, permet le maintien de la lysogénie, il est clivé par le
produit du gène recA dont la synthèse est induite par les UV; de ce fait, le
maintien de la lysogénie n’est plus possible et la bactérie lysogène passe
en cycle lytique.
Le passage de 37 °C à 42 °C a le même effet, dans des bactéries lysogénisées
par λts, porteur de la mutation CIts, que l’induction lytique des UV
chez une bactérie lysogénisée par un λ sauvage.
– Après empaquetage in vitro du phage λts/cos2, on infecte, avec une
multiplicité d’infection égale à 0,01, des bactéries d’une souche B, de
phénotype [F– ; ilv–; pro–; trp–; strR, recA+], dont les mutations sont
localisées sur la carte de coli (fig. 12.11).
On observe alors les résultats suivants :
b. Les bactéries infectées sont diluées et étalées sur boîtes (M0 + ile + val
+ trp + pro), on observe quelques plages troubles.
c. Les bactéries présentes dans ces plages troubles sont réisolées en clones
indépendants.
Plusieurs de ces clones, nommés B1, sont mis en culture à
37 °C, puis passés à 42 °C, on observe une induction lytique avec production
de particules phagiques.
d. On effectue un dosage de sodA dans un extrait protéique de B1, la dose
est double de celle présente chez la SSR (le dosage précis réalisé chez le
transformant A2 avait donné une valeur quatre fois supérieure).
Interprétez ces trois résultats (b, c, d) en montrant leur cohérence avec le
protocole suivi (a) dont vous justifierez, brièvement, les étapes et concluez.
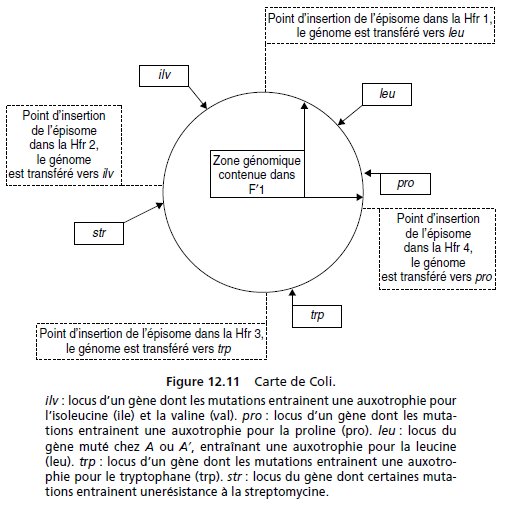
5. Quatre conjugaisons, interrompues après 40 min, sont réalisées en
parallèle entre des bactéries d’une colonie B1 et une des quatre Hfr de
phénotype sauvage (fig. 12.11).
On étale les conjugants sur un milieu M0
additionné de certains acides aminés et de str (tabl. 12.4, colonne 2).
Les colonies ainsi obtenues sont repiquées sur un même milieu, à 37 °C et
à 42 °C afin de tester leur capacité de pousser à cette température; le
tableau 12.4 donne les pourcentages des colonies repiquées poussant
à 37 °C et à 42 °C.
Donnez une interprétation cartographique précise et justifiée de ces résultats,
en discutant, notamment, du mécanisme conduisant au phénotype
capable de pousser à 42 °C, et des différences quantitatives observées (des
schémas clairs et précis sont demandés).
6. La même étude que celle faite pour A2, a été réalisée en parallèle pour la
souche A3; elle donne des résultats comparables à toutes les étapes sauf la
dernière; aucune des colonies recombinantes issues de la conjugaison avec
l’une des Hfr ne se révèle capable de pousser à 42° C.
Interprétez ce résultat, en montrant sa cohérence avec vos conclusions de
la question 3.
1. Les mutants sont létaux et ne peuvent être obtenus par un crible direct (problème 12.6).
Un gène est le plus souvent identifié par ses mutations et le(s) phénotype(s) mutant(s) qui en
résulte(nt).
Les croisements entre mutants et SSR sont un moyen de cartographier les mutations
et, partant, les gènes qui les portent.
En l’absence de mutants, il est donc impossible de cartographier des gènes dont on connaît
pourtant l’existence par la mise en évidence des activités enzymatiques qu’ils gouvernent.
2. a Ne pouvant pas isoler de mutants des gènes sodA ou sodB, on se propose de les cloner
dans un plasmide (ou, ici, un cosmide) et d’identifier les cellules transformées par un plasmide
recombinant porteur du gène sodA ou sodB par le fait que ces transformées devraient
présenter une activité superoxydes dismutase nettement accrue (problème 12.6).
NB : Ce protocole n’est donc possible que parce qu’on connaît la fonction des gènes et qu’on
sait doser leur produit.
Il est préférable de prendre une réceptrice (recA–) afin d’éviter les recombinaisons homologues
au locus sod, ce qui entraînerait la possibilité d’avoir des recombinants sauvages sans
surexpression; le gène sauvage n’étant plus plasmidique.
2. b Dans la figure 12.8 l’activité NADH déhydrogénase est un témoin interne permettant de
juger de la surexpression de l’activité sod dans l’extrait protéique.
La transformée A1 n’est
pas une transformée intéressante car il n’y a pas de surexpression de l’activité sod par rapport
à l’activité NADH déhydrogénase; en revanche, les transformées A2 et A3 présentent
respectivement une surexpression de l’activité sodA ou sodB par rapport à l’activité témoin
NADH déhydrogénase, et semblent a priori avoir acquis un cosmide porteur du gène sodA
ou du gène sodB.
3. Le transformant A3 surexprime le gène sodB et est [leu+].
Une interprétation possible
serait de considérer que le gène sodB est proche du gène leu, et que le cosmide contient la
séquence sauvage de ces deux gènes, mais ce n’est pas le cas :
– en effet, si c’était le cas, la séquence clonée dans pCOS3 aurait une carte de restriction
identique à la partie de la séquence sauvage de cette région contenue dans F′1 (fig. 12.11);
– par ailleurs, le lysat de A3 contiendrait des phages transducteurs porteurs de la séquence
sauvage leu+ (supposée être clonée avec sodB) et serait capable de transduire cette
séquence à la réceptrice A′ afin d’obtenir des transduites [leu+], ce qui n’est pas, non plus,
le cas.
Le gène sodB n’est donc pas localisé à proximité du gène leu et la séquence clonée dans
pCOS3 contient sans doute, outre le gène sodB, une séquence sauvage dont la surexpression
a un effet suppresseur sur l’effet de la mutation leu– de la souche A.
Le mutant A (leu–) possède pourtant cette séquence « suppresseur » qui a été coclonée avec
sodB dans pCOS3.
Mais chez A3, le « suppresseur » est cloné dans un vecteur multicopie et
il est surexprimé, il peut avoir alors l’effet suppresseur qu’il ne pouvait avoir en simple copie
chez A.