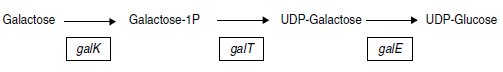
L’opéron galactose est constitué de trois gènes galE, galT et galK, transcrits
dans cet ordre, et codant respectivement pour une épimérase, une
transférase et une kinase, dans la chaîne métabolique suivante :
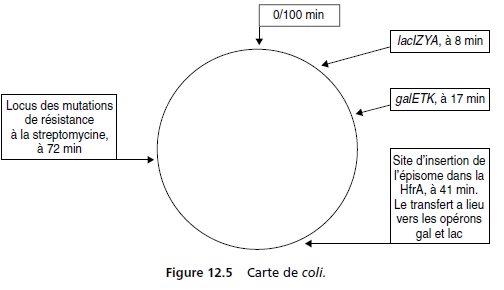
On dispose d’une souche HfrA de coli, mutée dans le gène galE de
l’opéron galactose (fig. 12.5), et sauvage pour l’opéron lactose.
Étalée sur milieu complet additionné de galactose (contenant une autre
source de carbone), la souche A ne peut pousser en raison de l’accumulation
d’UDP-galactose toxique pour la cellule; elle est donc sensible au
galactose, phénotype noté [GalS].
1. La souche A, étalée sur milieu complet additionné de lactose, se révèle
incapable d’y pousser. Interprétez ce résultat compte tenu de vos connaissances
en génétique et en métabolisme.
2. On étale une quantité importante de bactéries A issues d’une culture
fraîche sur des boîtes de milieu complet additionné de galactose (avec une
autre source de carbone), quelques colonies apparaissent, notée A1, A2,
etc. Interprétez ce résultat (plusieurs solutions possibles) en justifiant vos
réponses.
Ces colonies seraient-elles capables de pousser en présence de
lactose ?
3. Parallèlement, on étale une quantité importante de bactéries A issues
d’une culture fraîche sur des boîtes de milieu complet additionné de
lactose, quelques colonies apparaissent, notées Aa, Ab, etc.
Ces colonies
sont en quantité significativement supérieure au nombre de colonies A1,
A2, …, obtenues sur milieu complet additionné de galactose. Donnez une
interprétation qualitative et quantitative précise et justifiée de ce résultat
(plusieurs solutions possibles).
Ces colonies seraient-elles capables de
pousser en présence de galactose ?
4. On conjugue des bactéries des colonies A1, A2, et A3 avec une souche
(F– galE– ; strR), porteuse de la même mutation galE que la HfrA, et d’une
mutation de résistance à la streptomycine.
Puis on effectue des étalements
toutes les minutes sur un milieu complet additionné de galactose et de streptomycine, des colonies apparaissent à partir de la 24e minute. Interprétez
ce résultat après avoir justifié le protocole.
5. On conjugue des bactéries des colonies Aa, Ab, et Ac avec la souche
(F– galE–; strR), porteuse de la même mutation galE que la HfrA et d’une
mutation de résistance à la streptomycine.
Puis on effectue des étalements
toutes les minutes sur un milieu complet additionné de lactose et de
streptomycine :
– dans la conjugaison avec la HfrAa des colonies apparaissent à partir de
la 24e minute;
– dans la conjugaison avec la HfrAb des colonies apparaissent à partir de
la 33e minute;
– dans la conjugaison avec la HfrAc des colonies apparaissent à partir de
la 12e minute.
Interprétez ce résultat et montrez sa cohérence avec l’interprétation de
ceux de la question 3.
Solution
1. Le gène Z de l’opéron lactose code pour la β-galactosidase capable de cliver le lactose en
glucose et galactose; de ce fait la souche A va accumuler de l’UDP-Gal formé à partir du
galactose issu du lactose, ce qui conduira à la létalité.
Les mutants galE– sont donc de double phénotype [GalS, LacS], les mutations galE– ont un
effet pléiotrope du fait de l’interconnexion entre le métabolisme du galactose et celui du
lactose.
2. Ce sont des révertants, insensibles au galactose [GalR], ce qui ne signifie nullement qu’ils
peuvent métaboliser ce sucre.
Ils peuvent, par exemple, être [Gal–] par mutation dans le
gène galT, ce qui les rend insensibles au galactose puisqu’ils ne peuvent former d’UDP-Gal.
Comme pour tout révertant, plusieurs interprétations fonctionnelles sont possibles : révertant
vrai, suppresseur extragénique (par exemple, de non-sens), suppresseur intragénique (par
exemple, de décalage du cadre), deuxième mutation au site de la première, ou suppresseur
physiologique dont l’effet métabolique abolit l’effet de la mutation originelle; c’est le cas de
mutants galT ou galK où la formation d’UDP-Gal est bloquée.
Ces colonies A1, A2, sont a priori capables de pousser en présence de lactose puisqu’elles
sont devenues insensibles au galactose; le phénotype de sensibilité au lactose étant une
conséquence secondaire de la sensibilité au galactose.
3. Ce sont des révertants [LacR], insensibles au lactose.
Il y a parmi eux tous les révertants
[GalR] qui, de ce fait, deviennent aussi [LacR], mais il y a également des mutants [Lac–],
par mutation dans Z, qui deviennent insensibles au lactose puisqu’ils ne peuvent le métaboliser
et faire apparaître le galactose permettant l’accumulation d’UDP-Gal.
Non seulement, sur milieu lactose, le nombre de cibles mutables pour avoir des révertants
[GalR] est plus grand, mais certaines cibles sont aussi plus « faciles » à muter car correspondant
à des pertes de fonction, comme Z–; c’est pourquoi on obtient plus de colonies révertantes
sur ce milieu.
Evidemment les colonies Aa, Ab qui sont [lac–] ne peuvent pas pousser
sur galactose car elles ne sont [galR] qu’en raison de l’incapacité de le former à partir de lactose, mais demeurent [galS] en milieu avec galactose; le crible de sélection ayant ici isolé
des révertants sur l’effet second et non sur l’effet primaire de la mutation galE–.
4. A1, A2, A3 sont des révertants porteurs d’une séquence donnant la résistance au galactose
à 24 min de conjugaison, soit dans la région de l’opéron galETK (41 – 17 = 24 min !), ce
sont donc soit des révertants vrais, soit des révertants par suppresseur intragénique dans galE,
soit des mutants galT– ou galK–, suppresseurs physiologiques de galE– (voir problème 12.4).
5. Ce résultat illustre bien ce qui a été vu dans la question 3.
On a des révertants dans la
région gal, résistants au galactose et au lactose, et des révertants du type Hfrb, mutés dans la
région de l’opéron lacIZYA, sans doute Z–, de phénotype [lac–] et, de ce fait, [GalR].
Quant aux révertants de type Hfrc, ils sont porteurs d’un suppresseur de galE– à 41 – 12
= 29 min, donc hors de l’opéron galETK, ce qui laisse supposer que la mutation galE– est
d’un type admettant un autre suppresseur que les deux suppresseurs physiologiques possibles,
galT– et galK–, un troisième suppresseur, physiologique ou informationnel.
Problème 12.6
Chez E. coli, la biosynthèse de la lysine passe par l’acide diaminopimélique
(DAP), acide aminé indispensable à la constitution de la paroi
bactérienne; les mutants DAP– sont létaux et ne peuvent pousser que sur
un milieu additionné de DAP.
1. Proposez un protocole de sélection de mutants DAP–.
2. Un chercheur a obtenu de nombreux mutants DAP–, mais aucun n’a été
obtenu dans la dernière étape de sa biosynthèse, une transformation entre
l’isomère LL-Dap et l’isomère meso-DAP.
Pouvez-vous proposer quatre possibilités d’explication a priori de cette
absence de mutants ?
3. Les collègues biochimistes de ce chercheur lui révèlent l’existence de
l’activité DAP-isomérase catalysant cette réaction et le moyen de la
doser dans des extraits acellulaires.
Le chercheur se procure une banque plasmidique sauvage de E. coli, constituée de boîtes de colonies bactériennes
transformées par un plasmide pUN121 recombinant, porteur
d’un fragment d’ADN, issu d’une digestion partielle du génome sauvage
de coli.
Il met 300 clones en culture liquide, puis dose l’isomérase dans les
300 extraits acellulaires.
Trois colonies présentent une activité spécifique
trois fois supérieure à celle des autres colonies ou de la souche
sauvage.
Détaillez les diverses interprétations génétiques possibles de ce
résultat.
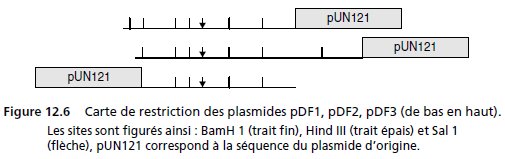
4. Le chercheur établit la carte de restriction des trois plasmides extraits de
ces trois colonies et nommés pDF1, pDF2 et pDF3 (fig. 12.6).
Qu’en conclut-il, d’un point de vue fonctionnel et cartographique ?
5. Un transposon contenant le gène de résistance au chloramphénicol (CmR)
est inséré entre le site Sal 1 et le site Hind III du troisième plasmide, noté
alors pDF3/Cm.
Après transformation de bactéries sauvages, les bactéries résistantes au
chloramphénicol présentent une activité spécifique en isomérase égale à
celle d’une souche sauvage.
Qu’en conclut notre chercheur ?
6. Il linéarise le plasmide pDF3/Cm au site Sal 1 et transforme une souche
de coli, acceptant la transformation par de l’ADN linéaire.
Sur quel milieu
se prépare-t-il à récupérer des transformants [CmR] ?
7. Notre chercheur finit par mettre en évidence ce qu’il cherchait; les transformants
[CmR] poussent sur une réplique sur Mo + chloramphénicol !
Qu’en conclut-t-il ?
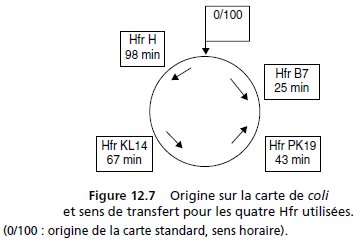
8. Puis il prépare sur ces transformants [CmR] un stock de phages P1 afin
de transduire quatre Hfr (H, B7, KL14 et PK19, fig. 12.7) en sélectionnant
pour la résistance au chloramphénicol.
Ces quatre Hfr sont mises à conjuguer avec une souche F– résistante à la
streptomycine, pendant 20 minutes avant interruption de la conjugaison et
étalement sur milieu complet + chloramphénicol + streptomycine. Seule la
conjugaison avec la Hfr KL14 donne des colonies sur la boîte d’étalement.
Quelle est sa conclusion ?
9. Profitant de l’abondance de son stock de phage transducteurs P1, le
chercheur transduit trois souches mutées respectivement dans le gène ilvA
(auxotrophe pour la valine et l’isoleucine), le gène rha (incapacité d’utiliser
le rhaminose) et le gène argE (auxotrophe pour l’arginine).
Que cherche-t-il ? Sur quels milieux va-t-il déposer ses cultures ?
10. Il teste les transductants obtenus pour leur résistance au chloramphénicol
et trouve que 30 % des transductants issus de la souche ilvA sont
résistants et qu’aucuns ne le sont pas parmi les transductants [arg+] ou
[rha+].
Que conclut-il ?
11. Le chercheur a-t-il fini par choisir parmi les quatre possibilités qu’il
s’était donné au départ. Pouvait-il espérer obtenir des mutants par le crible
proposé à la question 1 ?
Pourquoi ?
Ne vient-il pas de vous proposer un
crible adapté à ce type de mutants ?
12. Comment feriez-vous pour obtenir les autres mutants DAP–, sachant
que le temps vous est compté ?
Solution
1. On doit considérer un protocole en trois étapes :
– mutagenèse d’une culture en milieu supplémenté en DAP afin de permettre la multiplication
de mutants éventuels;
– enrichissement en milieu complet (mais sans DAP) avec pénicilline, afin de tuer les DAP+
et de préserver les DAP– qui restent en phase stationnaire et voient leur fréquence relative
augmenter, ce qui diminue le nombre de boîtes nécessaires à l’étape suivante;
– crible négatif en deux temps des mutants DAP, par étalement sur milieu supplémenté en
DAP et réplique des colonies sur milieu non supplémenté et identification sur la boîte mère
des mutants DAP n’ayant pas poussé sur la réplique.
2. Les quatre explications sont :
– hypothèse 1. Equilibre chimique ne nécessitant pas d’isomérase (cas connu, αD et βD
glucose);
– hypothèse 2. Isomérase nécessaire mais plusieurs isozymes existent, codée chacune par un
gène différent; la probabilité de toucher simultanément tous ces gènes est nulle;
– hypothèse
3. Isomérase nécessaire et unique mais indispensable dans une autre voie, la
mutation est alors létale car l’apport de DAP ne permet pas à cette autre voie de
fonctionner;
– hypothèse 4.
Mutagenèse et nombre de mutants criblés insuffisants (ce qu’on ne peut
jamais exclure a priori).
3. L’activité isomérase existe et est dosable, ce qui exclut l’hypothèse 1.
La banque plasmidique renfermant des fragments de digestion partielle est susceptible de
contenir une copie fonctionnelle de tous les gènes, y compris ceux renfermant un ou
plusieurs sites de restriction pour l’enzyme utilisée.
Les plasmides étant en général présents en plusieurs copies dans la cellule, les trois clones
identifiés par la surproduction d’isomérase sont susceptibles de renfermer la séquence du (ou
d’un) gène DAP.
Mais on pourrait aussi imaginer que le gène cloné est celui d’un activateur du (d’un)
gène DAP et que la surproduction de cet activateur permet la surproduction de l’enzyme.
4. L’alignement des cartes de restriction indique que les trois plasmides clonés semblent
porter une même séquence dans leur partie commune BBSHB, contenant au moins un
gène DAP.
Mais cela ne permet pas de savoir s’il existe un seul ou plusieurs gènes DAP, aussi
bien dans le génome que dans la séquence clonée, et si ce gène DAP est un gène de structure
de l’enzyme ou un gène régulateur.
5. Les bactéries résistantes ont acquis le plasmide recombinant pDF3/Cm.
L’activité isomérase résiduelle étant celle d’un sauvage non transformé, on peut conclure
qu’elle résulte de l’expression du chromosome et non de celle du plasmide, car il y aurait
alors surexpression.
L’absence de surproduction d’isomérase prouve que le plasmide pDF3
n’était porteur que d’un seul transgène et que celui-ci a été inactivé par l’insertion du transposon
(apportant la résistance).
Remarque. Le transposon est employé comme agent mutagène, et le gène de résistance
dont il est porteur est utilisé comme marqueur de sélection des transformants.
Mais ce résultat ne permet pas de statuer sur la fonction du transgène cloné dans
pDF3, gène DAP de structure ou gène régulateur.
6. La linéarisation du plasmide lui fait perdre le statut de réplicon et les bactéries qui, après
transformation, sont de phénotype [CmR] ont obligatoirement intégré le transposon et ses
séquences adjacentes dans son ADN chromosomique.
On sait que l’intégration d’une molécule d’ADN linéaire est ciblée par recombinaison homologue
avec les séquences homologues d’une de ses extrémités.
La linéarisation au site Sal1
cible l’insertion de l’ADN plasmidique dans le gène DAP cloné dans pDF3 (ou le gène de
l’activateur !) qui sera alors inactivé (fig. 12.3 ou 12.14).
Comme l’une des hypothèses à l’absence de mutants DAP est que l’inactivation de ce gène
pourrait être létale, le milieu de sélection des transformants doit contenir du DAP et du chloramphénicol
(pour la sélection des recombinants d’insertion).
7. Si le recombinant pousse sans apport de DAP, c’est qu’il est capable d’en produire, malgré
l’inactivation du gène cloné dans pDF3. Cette perte de fonction n’est donc pas létale, ce qui
exclut à présent l’hypothèse 3.
Plusieurs gènes redondants ayant même fonction isomérase,
l’hypothèse 2 semble la plus cohérente (à moins qu’il y ait plusieurs activateurs différents
mais isoactifs, cas peu vraisemblable).
De ce fait, l’hypothèse 4 est exclue, il était impossible, avec le crible utilisé, d’obtenir un
mutant DAP, car il aurait fallu simultanément et indépendamment toucher plusieurs gènes,
événement de probabilité nulle.
8. On sélectionne des recombinants [CmR] sachant que la séquence de résistance est insérée
dans un gène DAP.
En localisant la séquence de résistance dans les 20 minutes après
l’origine de transfert de KL14, soit entre 67 et 87 min, on localise en même temps ce
gène DAP.
Remarque. Le gène de sélection du plasmide et des mutants d’inactivation sert aussi
de marqueur de cartographie.
9. Les marqueurs d’auxotrophie choisis sont évidemment dans la zone 67-87 min et on
cherche à localiser plus finement le gène DAP en mettant en évidence une éventuelle
cotransduction entre DAP, c’est-à-dire le phénotype [CmR], et l’une des séquences sauvages
ilv+, rha+ ou arg+.
Les milieux d’étalement contiendront, selon les cas, de la valine, de l’isoleucine (recombinants arg+), de l’arginine (recombinants ilv+) ou de la valine, de l’isoleucine, de l’arginine et
du rhamnose (recombinants rha+).
La résistance pour le chloramphénicol, c’est-à-dire la cotransduction du gène DAP, sera
testée par réplique sur un même milieu additionné de l’antibiotique.
Remarque.
On aurait pu imaginer tester directement la croissance sur ces trois
milieux en présence de chloramphénicol, mais l’absence de colonies aurait pu être
alors interprétée soit comme l’absence de cotransduction, soit comme une erreur de
l’expérimentateur dans la réalisation du protocole de transduction.
L’étalement sur
une première boîte, en fournissant des colonies, prouve que l’expérimentateur a
convenablement travaillé et exclut toute ambiguïté dans l’interprétation d’un résultat
négatif sur la réplique.
10. Le gène DAP est cotransduit (30 %) avec le gène ilvA, ce qui permet, connaissant la carte
de coli, une localisation assez fine.
11.
Si une fonction biologique est assurée par des gènes redondants (comme DAP ici), il est
impossible d’avoir des mutants dont le phénotype diffère de sauvage par l’absence de cette
fonction car on ne peut obtenir, par une seule mutagenèse, des mutants de tous ces gènes à la
fois, mutants qui seraient par ailleurs létaux; quant aux mutants simples, ils gardent un
phénotype sauvage !
L’astuce réside ici dans le détour qui consiste à cloner l’un de ces gènes dans un vecteur
d’expression, en sélectionnant un clone surproducteur de l’activité, puis en inactivant le gène
cloné et en ciblant l’intégration du vecteur de clonage dans le gène chromosomique, le
recombinant étant identifiable par un gène de sélection positive (ici la résistance au chloramphénicol).
On obtient alors un mutant du gène, ce qui était impossible par mutagenèse
directe.
12. Ce protocole est assez long, notamment le dosage de la banque génomique, or on peut
imaginer que les autres gènes DAP ont une assez bonne homologie avec celui cloné dans
pDF3.
On peut alors envisager d’utiliser la séquence de ce gène DAP comme sonde pour trier une
banque d’ADN génomique.
Les clones ainsi identifiés peuvent être séquencés (opération
facile et rapide aujourd’hui) afin de comparer les séquences avec DAP.
La localisation génomique des séquences identifiées peut suivre le même protocole.