4. On obtient évidemment des révertants [his+], soient des révertants vrais, soit des révertants
par deuxième mutation au site de la mutation directe, soit des révertants par suppresseur
intragénique, ou suppresseur informationnel ou suppresseur physiologique (s’il en existe un
potentiel).
Le révertant K1-a est aussi [pur+], ce qui semble indiquer que le suppresseur pourrait être un
suppresseur informationnel opérant à la fois sur la mutation his1 et la mutation pur–, qui
seraient alors toutes deux des mutations stop de même nature (l’effet Wobble n’ayant pas été
vu dans les rappels, il n’est pas attendu plus de précision dans la réponse).
5. On sait, par les conjugaisons interrompues entre K et F1, que la séquence his1+ entre à
i minutes de l’origine de transfert.
Le croisement entre F1 et le révertant Hfr K1-a permettra d’obtenir des recombinants [his+]
à i min de conjugaison si le révertant est un révertant vrai ou un révertant muté au site de la
mutation directe ou un révertant avec un suppresseur très proche de celle-ci.
Mais si le révertant
est porteur d’un suppresseur éloigné de la mutation directe his1, il faudra attendre
l’entrée de ce suppresseur pour avoir des recombinants [his+], puisque la F1 comme la Hfr
sont porteuses de la même séquence his1 mutée.
Comme on obtient des recombinants [his+] dès la minute (i – 6), on peut conclure que le
révertant K1 est bien porteur d’un suppresseur localisé 6 minutes en amont de la mutation
his1 (dans le sens du transfert) et 6 minutes en aval, sur la carte de coli.
6. La Hfr K1-a est porteuse des séquences his1– et his2+, ainsi que du suppresseur de his1–,
localisé six minutes en amont (dans le sens du transfert).
Si on n’obtient aucun recombinant avant i minutes, alors que le suppresseur est transféré six
minutes plus tôt, on peut conclure que le suppresseur de his1– n’a aucun effet sur his2–, ce
qui est cohérent avec l’hypothèse d’un suppresseur informationnel, dont on sait qu’il est
allèle spécifique (his1– et pas his2–) et gène non spécifique (his1– et thr–).
On peut en conclure que :
– soit la mutation his2– n’est pas une mutation stop;
– soit, éventuellement, qu’elle est une mutation stop différente de his1– et ne pouvant pas
être corrigée par le suppresseur de his1–;
– soit qu’elle est une mutation stop identique à his1–, mais que l’acide aminé, apporté par
l’ARN-t suppresseur, ne restaure pas la fonctionnalité de la chaîne peptidique.
Il faut donc attendre i minutes pour que le transfert de la séquence his2+ de la Hfr K1-a puisse
permettre l’obtention de recombinants [his+].
Problème 12.4
On dispose, chez E. coli, d’une souche Hfr, porteuse d’une mutation galU1
la rendant sensible au galactose.
Ce mutant, dont le phénotype est noté
[GalS], est tué sur milieu complet additionné de galactose parce qu’il accumule
des quantités toxiques d’UDP-Gal et d’UDP-Glu.
On doit donc lui
fournir une autre source de carbone dont la voie de métabolisation évite
une telle accumulation. Le phénotype sauvage est résistant au galactose et
est noté [GalR].
Après culture de ce mutant Hfr galU1, on a fait des étalements sur un
milieu complet additionné de Galactose, et on a obtenu quelques colonies
à l’origine de souches R, dont se propose de faire l’analyse génétique.
1. Quel est le phénotype de telles souches ?
Leur génotype ?
2. On croise les diverses souches R avec la souche S (F–; galU1, strr), où
strr est une mutation de résistance à la streptomycine et, selon un protocole
de conjugaison interrompue, on effectue des prélèvements pour tester la
présence de recombinants [GalR].
Selon la souche R utilisée, le temps minimal de croisement pour obtenir
des recombinants sur une boîte d’étalement est différent, ce qui permet de
définir trois types de souches :
– les souches de type R1, pour lequel ce temps est de 2 minutes;
– les souches de type R2, pour lequel il est de 25 minutes;
– les souches de type R3, pour lequel il est de 45 minutes.
La conjugaison interrompue entre une Hfr sauvage (de même origine de
transfert que la Hfr galU1) et la souche S donne des colonies sur le milieu
d’étalement à 25 minutes.
a. Quel est le milieu d’étalement permettant d’identifier les recombinants
[GalR] ?
b. Proposez un génotype pour chacun des types de souches R1, R2 et R3.
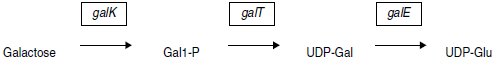
3. On dispose de plasmides porteurs d’un gène de résistance à la kanamycine
(kanR) et d’un des gènes de l’opéron galactose gouvernant la métabolisation
du galactose, dont on rappelle ci-dessous la séquence :
Ces plasmides sont dénommés pgalE, pgalK et pgalT, selon le gène de
l’opéron gal qui y est cloné (le gène cloné est fonctionnel); un quatrième
plasmide, noté pgalEKT est porteur de la totalité de l’opéron.
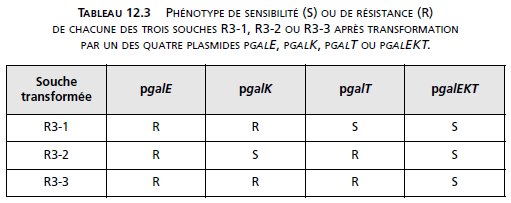
On transforme, par ces plasmides, trois souches de type R3, nommées R3-1, R3-2
et R3-3, et on sélectionne les transformants [kanR] que l’on teste pour leur
sensibilité au galactose (tabl. 12.3).
a. Quelle fonction peut être affectée dans chacune des souches de type R3-1,
R3-2 et R3-3 ? Justifiez votre réponse en précisant la cause du phénotype
de résistance au galactose et de restauration du phénotype sensible chez les
transformées.
b. La souche R3-3 est transformée avec un plasmide porteur d’un gène de
résistance au chloramphénicol (CmR) et d’un suppresseur d’ambre, les
transformants [CmR] sont [GalS].
Donnez une interprétation génétique et moléculaire cohérente des deux
observations concernant cette souche R3-3, sachant, par ailleurs, que le
gène galE est le premier des gènes transcris de l’opéron galactose.
4. Sachant que la métabolisation du galactose comme source de carbone
est inductible par le galactose lui-même, à quels types de mutants pourraient
correspondre les souches de type R1 ?
Quelle est la localisation de
l’opéron galactose ?
5. Quel test génétique simple (aucune « cuisine » n’est demandée) devrait
permettre de choisir entre les deux hypothèses formulées précédemment ?
Solution
1. Les souches R sont de phénotype [galR] et sont des révertants, soit des révertants vrais,
soit, plus probablement, des révertants où la mutation directe (originelle) voit son effet
supprimé par celui d’une nouvelle mutation à effet suppresseur, nommée sua (pour suppresseur
actif, l’allèle sauvage étant noté sui); la mutation sua peut être intra ou extragénique par
rapport à la mutation directe.
Le génotype du révertant peut donc être galU1+ ou (galU1; sua).
2. a On étale sur milieu complet additionné de galactose (sélection de recombinant résistants)
+ streptomycine (contre sélection des donatrices).
2. b Le croisement entre la Hfr sauvage et la réceptrice galU1 permet de localiser la mutation
galU1 à 25 min de conjugaison.
La Hfr révertante R1, après 2 min de conjugaison, donne à la réceptrice galU1 une séquence
lui conférant le phénotype [galR]; elle possède donc à cet endroit, éloigné de galU1 de
23 min (donc extragénique), une mutation suppresseur.
De la même façon, la révertante R3 possède un suppresseur localisé à 45 min de l’origine de
transfert.
La révertante R2 confère à la réceptrice S (galU1) un phénotype [galR] en lui transférant une
séquence localisée à 25 min, comme galU1.
Du fait de la grossièreté de la cartographie, par
conjugaison il est impossible de savoir si R2 est un révertant vrai ou un révertant par suppresseur
intra ou même extragénique.
Remarque.
La mise en évidence d’un suppresseur, en génétique eucaryote, est entreprise
par l’étude de la méiose chez le diploïde issu du croisement entre le révertant et
la souche sauvage, afin d’isoler d’éventuels gamètes recombinants, où la mutation
directe aurait été séparée de son suppresseur.
En génétique bactérienne, où il n’y a ni diploïde ni méiose, on croise le révertant Hfr
avec une réceptrice mutée au même site de mutation directe que la Hfr, à partir de
laquelle on a isolé les révertants; un éventuel suppresseur sera mis en évidence par
cartographie puisque c’est la seule séquence de la Hfr révertante pouvant donner des
réceptrices sauvages par conjugaison.
La seule ambiguïté demeurera pour le cas où la
séquence Hfr, donnant des recombinants sauvages, serait localisée très près de la
mutation directe.
3. a Le plasmide pgalT apporte un gène actif galT au révertant R3-1, ce qui le rend [galS],
alors que l’apport de galE ou galK ne changent pas son phénotype [galR].
Il est facile de
conclure que R3 est un révertant [galR] par mutation dans son gène galT, ce qui entraîne
l’impossibilité de transformer Gal-1P en UDP-Gal, d’où le phénotype [galR]. Le phénotype
[galS] mutant est restauré par l’apport plasmidique de galT.
Bien évidemment, l’apport
de tout l’opéron a le même effet que l’apport de gal uniquement.
De la même façon, on peut conclure que le révertant R3-2 est muté dans galK; l’apport de
galK restaure le phénotype muté [galS] en permettant à la transformée de phosphoryler le
galactose en Gal1-P.
Le révertant R3-3 est muté dans l’opéron gal puisque le plasmide pgalEKT restaure la sensibilité,
mais n’est pas muté dans un seul gène de cet opéron puisqu’aucun des trois gènes
apporté isolément ne restaure la sensibilité; R3-3 est muté au moins dans le gène GalT et le
gène galK ou galE de l’opéron puisque la résistance suppose l’absence d’UDP-Gal, et que la
résistance n’est restaurée qu’avec l’apport de ces deux gènes.
La mutation peut être unique s’il s’agit d’une délétion chevauchant ces deux gènes.
Remarque. On a ici un exemple typique de suppresseur physiologique.
La perte de
fonction d’un gène galK ou galT empêche certes toute croissance sur galactose mais
a, en contre-partie, l’effet positif de sauver les mutants galU1 de la toxicité du galactose.
Ces mutants peuvent pousser, pour peu qu’ils trouvent dans le milieu une autre
source de carbone.
Aussi il convient toujours de se méfier des jugements hâtifs qui consisteraient à considérer
qu’une perte de fonction ne peut avoir qu’un effet négatif…
La conséquence
positive ou négative de l’effet d’une mutation ne peut se juger qu’en fonction du
contexte génétique et du milieu !
3. b Les transformants [CmR] ont acquis le plasmide porteur du suppresseur d’ambre et
retrouvent la sensibilité.
Celle-ci ne peut résulter que de l’action du suppresseur d’ambre, ce
qui prouve que l’opéron gal du révertant n’est pas muté par délétion mais par mutation ambre
(non-sens UAG).
Comme au moins deux fonctions (galK et galT ou galT et galE) sont absentes chez le révertant
R3, peut-on supposer que les deux gènes soient mutés par un stop UAG ?
La probabilité
d’un tel événement quadruple, deux mutations indépendantes et de même nature UAG, est
trop faible pour qu’une telle hypothèse puisse être crédible.
L’indication de l’ordre des gènes permet de faire une autre hypothèse plus crédible car
supposant que le révertant R3-3 serait porteur d’une seule mutation UAG, dans un seul gène,
avec un effet polaire bloquant l’expression des gènes en aval.
En effet, chez la bactérie, la traduction étant couplée à la transcription, l’arrêt de la traduction
sur une longueur suffisante de l’ARN-m entraîne l’arrêt de la transcription, dont la poursuite
dépend de façon critique de l’occupation de l’ARN par des ribosomes.
On peut donc considérer que R3-3 peut être porteur d’une mutation ambre soit dans le
gène galE (les gènes galE, galT et galK ne sont pas exprimés), soit dans le gène T (les gènes
galT et galK ne sont pas exprimés).
4. L’opéron galactose touché dans les révertants de type R3 est localisé à 45 min de l’origine
de transfert; les révertants de type R1 ne sont donc pas touchés dans l’opéron galactose mais
pourraient être touchés dans un gène de régulation de cet opéron :
– soit par perte de fonction, s’il s’agit d’un activateur, l’absence de transcription de l’opéron
gal conduisant au phénotype de résistance;
– soit par une mutation d’incapacité de lier le galactose (dont le rôle est inducteur) s’il s’agit
d’un répresseur.
On aurait alors un mutant surréprimé conduisant au phénotype de
résistance.
Ce type de mutation correspond aux mutations IS du gène I du répresseur régulant la transcription
de l’opéron lactose.
5. On sait qu’une mutation de perte de fonction dans le gène d’un activateur aurait un effet
« récessif » par rapport à son allèle sauvage, tandis qu’une mutation de surrépression (de
type IS) a un effet dominant sur celui de sa séquence sauvage.
On peut envisager, pour départager
les deux hypothèses, de réaliser un tel test de dominance en observant le phénotype
d’un diploïde partiel obtenu chez R1 après transformation avec un plasmide ou un épisome
(sexduction) apportant la séquence sauvage localisée autour des 2 min de conjugaison.