On dispose, chez coli, des souches A, B, C et D de génotype :
A : (cysB–; tyr1–; tyr2+; strR); B : (cysB–; tyr1+; tyr2–; strR);
C : (cysB+; tyr1–; tyr2+; strR); D : (cysB+; tyr1+; tyr2–; strR).
où cysB, tyr1 et tyr2 sont des mutations ponctuelles respectivement
responsables d’une auxotrophie pour la cystéine ou pour la tyrosine. strR
étant une mutation de résistance à la streptomycine.
1. Des expériences de conjugaisons interrompues, avec 15 secondes de
précision, entre une Hfr sauvage et des réceptrices tyr1 ou tyr2 donnent un
temps de transfert égal pour tyr1+ et tyr2+, à 30 secondes après cysB+,
plusieurs dizaines de minutes avant le site str.
a. Quelles ont été les réceptrices utilisées ?
b. Quels ont été les milieux de cocultures et d’étalement ?
c. Qu’en concluez-vous pour tyr1 et tyr 2 sur le plan cartographique ?
Sur
le plan fonctionnel ?
Justifiez vos réponses de manière claire et concise. (Quelques lignes suffisent,
à chaque fois, ne perdez pas de temps sur cette question.)
2. Par un test trois points on a montré que l’ordre était : cysB-tyr2-tyr1.
Vous reconstituerez le protocole de ce test trois points, réalisé par transduction,
sachant qu’on disposait de phage transducteur P1 et des quatre
souches de E. coli, A, B, C et D.
a. Quels sont les croisements effectués ?
Avec quels lysats à partir de
quelles donatrices ?
Sur quelles réceptrices ?
Quels sont les recombinants
sélectionnés ?
Sur quels milieux ?
b. Quels sont les résultats possibles ? quel est le résultat obtenu ?
Quelle
est l’argumentation permettant de justifier la conclusion à partir du résultat
obtenu ?
On demande des réponses claires, concises, argumentées, accompagnées
des schémas utiles.
3. Les souches A et B sont transformées par un plasmide PK, porteur d’un
gène de résistance à la kanamycine et d’un fragment de génome sauvage
de coli contenant, entre autres, la séquence cysB+.
Les colonies [kanR], issues de la transformation de A ou B par PK sont,
[cys+, tyr+]. Concluez.
4. Un transposon est une séquence d’ADN capable, par elle-même, de se
déplacer pour s’insérer (ou de se dupliquer pour insérer une copie) plus ou
moins aléatoirement en un autre site du même réplicon ou d’un autre
réplicon.
L’insertion d’un transposon dans la séquence d’un gène a souvent un effet destructeur sur le message de ce gène qui ne peut plus être
exprimé; elle entraîne donc une perte de fonction du gène.
La plupart des transposons bactériens sont porteurs d’un gène conférant la
résistance à un antibiotique.
De nombreux plasmides, porteurs d’un gène
de résistance à un antibiotique, sont porteurs du transposon où ce gène est
inclus.
On soumet le plasmide PK à l’action d’un transposon porteur d’un gène de
résistance à l’ampicilline, puis on sélectionne un plasmide recombinant
conférant la double résistance à la kanamycine et à l’ampicilline; ce plasmide
est appelé PKA.
• Les colonies [kanR; ampR] issues de la souche A transformée par PKA
sont [cys+, tyr+].
• Les colonies [kanR; ampR] issues de la souche B transformée par PKA
sont [cys+, tyr–].
Qu’en concluez-vous pour le plasmide PKA et pour les mutations tyr1
et tyr2 ?
Solution
1. a Celles qui, ayant une séquence mutée, peuvent être sélectionnées comme recombinant
sauvage après croisement avec la Hfr, soit, A pour cys et tyr1, et B pour cys et tyr2.
1. b Le croisement (coculture) se fait en milieu permettant la pousse des deux souches, soit
Mo + cys + tyr.
Le milieu d’étalement est destiné à la sélection des recombinants; il est donc
additionné de streptomycine pour bloquer la pousse des donatrices et de tyr pour évaluer le
temps de transfert de cys+, ou de cys pour évaluer le temps de transfert de tyr1+ ou tyr2+.
1. c Les deux mutations tyr1 et tyr2 sont localisées au même endroit, à la précision près de la
cartographie par conjugaison interrompue, soit ici 15 secondes ou 10 000 pb.
Bien évidemment, la proximité des mutations ne signifie nullement qu’elles touchent le
même gène; c’est possible, mais elles peuvent aussi toucher deux gènes proches impliqués
tous deux dans la biosynthèse de la tyrosine.
2. Il convient de faire des lysats transducteurs des souches C et D afin de transformer respectivement
les souches réceptrices B et A.
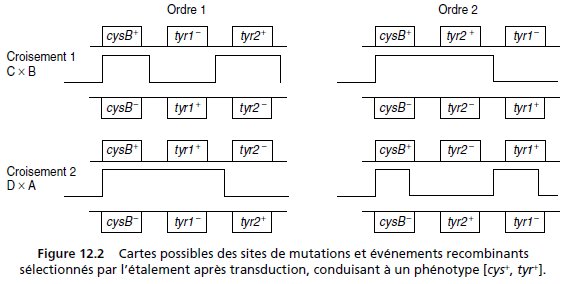
Il y a deux ordres possibles pour les trois sites cysB, tyr1 et tyr2 (fig. 12.2) puisqu’on sait que
cysB ne peut être central, étant transféré 30 secondes avant tyr1 ou tyr2.
Pour chacune des deux transductions les combinaisons alléliques sont différentes (fig. 12.2)
le phage apportant tyr1– et tyr2+, ou tyr1+ et tyr2–.
On étale une quantité connue de cellules sur une boîte de milieu minimum où seules les
recombinants sauvages [cys+, tyr+] de génotype (cysB+, tyr1+, tyr2+) peuvent pousser.
Si l’ordre 1 est l’ordre réel on attend, dans le croisement 1, une fréquence de recombinants
sauvages [cys+, tyr+] très inférieure à celle du croisement 2; car la formation de génotypes
sauvages y nécessite quatre événements de recombinaison (fig. 12.2) alors que deux sont
suffisants dans le croisement 2.
Si l’ordre réel est le 2, on attend le résultat contraire, c’est-à-dire celui qui a été observé, une
fréquence de recombinants sauvages très inférieure dans le croisement 2. Cet ordre est validé.
3. Si les transformées ayant reçu le plasmide PK, puisque résistantes à la kanamycine, sont
de phénotype [cys+; tyr+], c’est que le fragment génomique cloné dans PK est porteur des
séquences sauvages cysB+, tyr1+ et tyr2+.
Comme il s’agit, pour les trois mutations, d’un diploïde partiel, on peut conclure que les
mutations d’auxotrophie ont un effet récessif par rapport à leurs allèles sauvages respectifs.
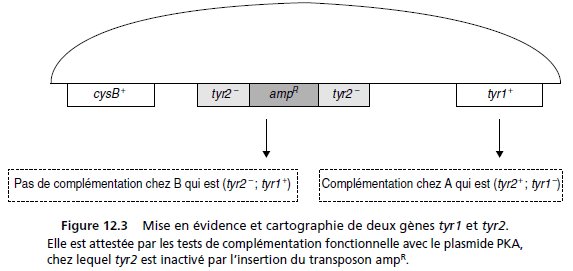
4. Il y a insertion du transposon dans PK et formation de PKA.
Le phénotype [cys+, tyr–] des colonies [kanR, ampR] permet de conclure que la réceptrice B
comme le plasmide PKA sont mutés dans le même gène.
Il s’agit de la mutation tyr2 chez B,
et de la « destruction » de la séquence de ce gène par insertion du transposon, chez PK, lors
de la formation de PKA.
La « destruction » d’un gène entraîne une perte de fonction et est, sans doute, récessive.
Le phénotype [cys+, tyr+] des colonies [kanR, ampR] permet de conclure qu’il y a complémentation
fonctionnelle et que la réceptrice A, porteuse de la mutation tyr1, n’est pas mutée
dans le même gène que le plasmide PKA, (sinon le phénotype serait [tyr–]) et que les mutations
tyr1 et tyr2 sont proches et touchent des gènes voisins, voire contigus (fig. 12.3).
Une souche F– de E. coli, mutée dans les deux gènes purF et pheS est de
phénotype [pur–; phe–], auxotrophe pour les purines et la phénylalanine.
À partir de cette souche on isole deux mutants [his–], auxotrophes pour
l’histidine et notés F1 et F2, les mutations étant respectivement notées his1
et his2.
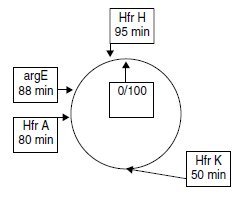
On dispose des trois souches Hfr H, A ou K, chacune délétée pour le
gène argE, situé à 88 min sur la carte de coli.
La Hfr H transfère ses gènes
dans le sens des aiguilles d’une montre, les deux autres Hfr dans le sens
trigonométrique.
1. On croise chacun des mutants F1 et F2 par la HfrH, A ou K.
On étale les conjugants après 50 minutes de croisements, après les avoir
séparés par vortex, sur un milieu Mo + pur + phe.
On obtient des colonies dans tous les cas.
a. Quelle est votre conclusion ? Justifiez vos réponses.
b. Justifiez le phénotype des Hfr.
2. On recommence les croisements entre la Hfr K et les souches F1 et F2.
On étale les conjugants après 30 minutes de croisement, après les avoir
séparés par vortex, sur un milieu Mo + his + pur.
Les colonies obtenues sur ces boîtes mères sont testées par réplique sur des
boîtes Mo + pur, 60 % y poussent, et sur des boîtes Mo + his, 40 % y
poussent.
On n’obtient pas de recombinants [phe+] dans le croisement entre la Hfr A
et les réceptrices F1 ou F2, après 50 minutes de croisement, mais on
obtient des recombinants [pur+] ou [his+].
Qu’en concluez-vous ? Justifiez vos réponses.
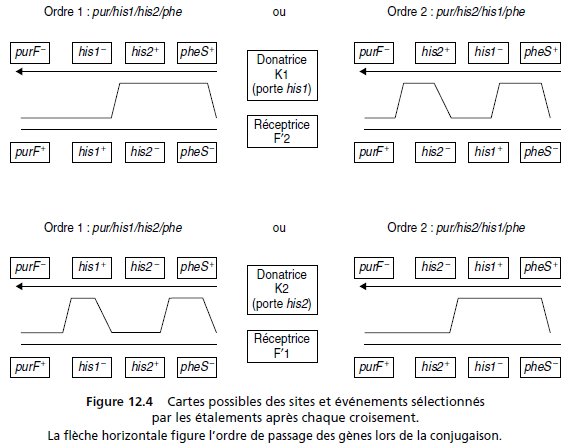
3. La conjugaison interrompue montre que les séquences his1+ et his2+
sont localisées au même endroit de la carte, i minutes après l’origine de transfert de K.
On sélectionne, parmi les colonies testées à la question
précédente, des colonies F′1 et des colonies F′2 de phénotype [pur+, his–,
phe–] afin de les croiser avec des Hfr dérivées de K, porteuses de la même
mutation d’auxotrophie aux purines que la souche F, et porteuses, soit de
la mutation his1, soit de la mutation his2.
Ces Hfr sont respectivement
notées K1 et K2.
• Le croisement K1 avec F′2 [pur+, his–, phe–] est réalisé, le titre de F′2
est de 108/ml, et on observe 225 colonies pour un étalement de 0,1 ml, sur
milieu Mo.
• Le croisement K2 avec F′1 [pur+, his–, phe–] est réalisé, le titre de F′1
est de 5 × 108/ml, et on observe 275 colonies pour un étalement de 0,1 ml,
sur milieu Mo.
Qu’en concluez-vous ? Justifiez vos réponses en les accompagnant de
schémas clairs et précis, et en justifiant les génotypes des souches utilisées
dans les croisements.
4. Une culture de la Hfr K1 est irradiée aux UV et on étale 108 cellules sur
une boîte de milieu Mo + pur + arg.
On obtient quelques colonies, dont
une, notée K1-a, que l’on étudie.
Répliquée sur un milieu Mo + arg, les cellules de la colonie K1-a se révèlent
capables d’y pousser.
Donnez une interprétation fonctionnelle simple à tous ces résultats.
5. On réalise une cinétique de conjugaison interrompue toutes les
30 secondes entre la Hfr K1-a et la réceptrice F1.
On obtient des recombinants
[his+] au temps (i – 6) minutes.
On rappelle que les mutations his1 et his 2 sont localisées à i minutes de
l’origine de transfert de K.
Donnez une interprétation cartographique et fonctionnelle simple à tous
ces résultats, et montrez leur cohérence avec vos conclusions de la
question 4.
6. On réalise une cinétique de conjugaison interrompue toutes les
30 secondes entre la Hfr K1-a et la réceptrice F2.
On obtient des recombinants
[his+] au temps i minutes.
Quelles précisions apportent ces résultats ?
Solution
1. a On recueille les réceptrices recombinantes de phénotype [his+]; les séquences mutées
his1 ou his2 y sont remplacées par leurs homologues sauvages his1+ ou his2+. Celles-ci sont
donc entrées entre 95 et 45 min avec la Hfr H (sens horaire), entre 50 et 0 min (sens trigo)
avec la Hfr K, et entre 80 et 30 min (sens trigo) avec la Hfr A.
Donc les séquences mutées
sont dans la partie commune de ces trois transferts, soit entre 30 et 45 min de la carte de coli.
b. La délétion du gène argE sert de marqueur de sélection des réceptrices et de contresélection
des donatrices; la séquence délétée de la Hfr n’est jamais transmise dans les 50 min
de croisement.
2. On sélectionne, après croisement de 30 min avec la Hfr K, des recombinants [phe+]; le fait
que 60 % d’entre eux soient aussi [his+] alors que 40 % d’entre eux sont aussi [pur+] montre
que les séquences his1+ ou his2+ sont plus proches du gène pheS (corecombinaison plus
fréquente, 60 %) que du gène purF; ces deux gènes purF et pheS étant localisés dans les
30 premières minutes de transfert de la HfrK.
L’ordre des marqueurs est donc purF-(his1/his2)-pheS, ou bien (his1/his2)-pheS-purF, avec
une distance his-pheS plus petite que la distance pheS-purF.
Le fait de ne pas obtenir de recombinants [phe+] avec la Hfr A alors qu’on en obtient avec la
Hfr K indique que le gène pheS est à plus de 30 min de l’origine de transfert de A, et à moins
de 30 min de l’origine de transfert de K; le gène pheS est donc le gène le plus distal de
l’origine de transfert de K ou de A.
L’ordre des marqueurs, dans le sens de l’ordre de transfert par la Hfr K est purF-(his1/his2)-
pheS, ce qui donne sur la carte standard de coli, l’ordre PheS-(his1 ou his2)-purF.
3. Les mutations his1 et his2 sont situées à i min de transfert de l’origine de K, et ne peuvent
être cartographiées l’une par rapport à l’autre par une simple conjugaison interrompue.
Deux ordres sont possibles (fig. 12.4) : purF-his1-his2-pheS ou purF-his2-his1-pheS dont le
choix dépendra des résultats des deux croisements réalisés en parallèles, et de l’étude de la fréquence des recombinants [pur+; his+; phe+] obtenus dans ces deux croisements, où pheS
sert de marqueur distal de recombinaison.
Si l’ordre 1 est correct, on attend plus de recombinants sauvages (en fréquence) dans le
premier croisement (fig. 12.4) que dans le second.
Si l’ordre 2 est correct, on attend le
résultat inverse.
On observe 225 recombinants dans le premier croisement, et 275 dans le second qui doivent
être rapportés à des titres respectifs de 108/ml et de 5 × 108/ml, ce qui donne des fréquences
de recombinants respectivement égales à 2,25 × 10–5 et 0,55 × 10–5 (dépôt de 0,1 ml contenant
respectivement 107 et 5 × 107 bactéries).
La fréquence des recombinants étant supérieure dans le premier croisement, l’ordre 1 est
validé.