



















Équilibre acide-base :
A – PRINCIPES FONDAMENTAUX :
1- Ion H+ :
L’organisme de l’adulte soumis à un régime alimentaire équilibré génère quotidiennement, dans son état de fonctionnement de base normal, une quantité de protons (H+) d’environ 50 à 100 mEq,
 alors que la production d’anhydride carbonique (CO2), susceptible par hydratation de créer elle aussi des protons est considérable (environ 15 000 mmol/jour).
alors que la production d’anhydride carbonique (CO2), susceptible par hydratation de créer elle aussi des protons est considérable (environ 15 000 mmol/jour).
Cet ion H+ est particulièrement important en biologie, notamment en raison du champ électrique majeur qui lui est associé, et qui le rend particulièrement réactif.
Cette propriété est en relation avec la taille relativement faible de l’ion hydronium (H3O+), la forme hydratée de l’ion H+, notamment par comparaison à la taille de cations comme Na+ et K+.
Dans l’environnement aqueux des liquides biologiques, en effet, le proton H+ se combine à l’eau, et il est essentiellement présent sous la forme H3O+.
Toutefois, par souci de simplification, nous parlons, dans la suite de ce texte et comme c’est très généralement le cas dans la littérature, de H+, en lieu et place de H3O+.
2- Concentration protonique : pH
De nombreuses enzymes très importantes sont hautement sensibles à leur environnement protonique.
Leur fonction s’altère de façon majeure lorsque cet environnement varie, alors que les liaisons de substances importantes comme, par exemple, certaines hormones, soit avec des protéines circulantes, soit avec leurs récepteurs membranaires, sont influencées par la concentration protonique du milieu dans lequel elles sont localisées.
Il n’est donc pas étonnant que la concentration de l’ion H+ doive être très étroitement contrôlée.
Ce contrôle permet, en conditions normales, le maintien d’une concentration protonique de 40 ± 5 nmol/L (10–9 mmol/L), même si un flux continuel de bases et d’acides s’ajoute aux liquides intra- et extracellulaires.
Il s’agit donc d’un équilibre homéostatique, d’un état stationnaire.
Il est également très important de prendre note de l’échelle de mesure de la concentration du proton dans les milieux biologiques (nmol/L), dont la concentration est très faible comparée à celle d’autres ions, comme le sodium, le potassium ou le chlore, qui sont mesurés en mmol/L, soit dans un ordre de grandeur très différent.
Au laboratoire, la concentration de H+ est mesurée avec une électrode de verre perméable seulement aux protons.
La diffusion de H+ provenant de l’échantillon sanguin vers l’électrode permet la génération d’une différence de potentiel électrique, proportionnelle au logarithme de la différence des concentrations de H+ de part et d’autre de la membrane de l’électrode, selon l’équation de Nernst.
Celle-ci veut que le potentiel électrique généré soit inversement proportionnel au logarithme de la concentration en protons de l’échantillon, donc à son logarithme négatif, ou à son antilogarithme, ou encore cologarithme, appelé pH (potentiel hydrogène).
Cet index est devenu l’expression usuelle de la concentration protonique en biologie et en médecine, soit :
pH = – log [H+] (équation 1)
Pour être tout à fait strict, l’électrode est directement sensible à l’activité de H+ et non à sa concentration, cette activité étant définie par les mouvements ioniques effectués au hasard de part et d’autre de la membrane de l’électrode de mesure.
Toutefois, l’activité ionique est fonction de la concentration de l’ion dans le milieu, via un coefficient appelé force ionique qui traduit les interactions entre les ions eux-mêmes, lesquelles sont directement proportionnelles à leur concentration.
Comme la dilution des protons en milieu aqueux est extrême sans grand risque d’erreur dans ce cas, activité et concentration sont considérées équivalentes.
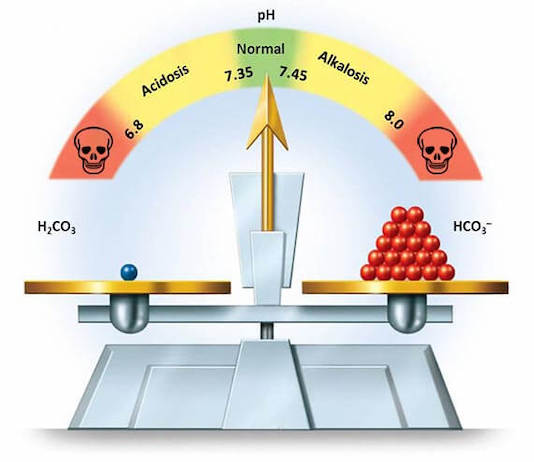
Si le pH normal est de 7,40 ± 0,05, que, en raison de l’échelle logarithmique inhérente à cette unité de mesure protonique, l’augmentation de [H+] lorsque la valeur du pH décroît, ou la diminution de [H+] quand le pH augmente, ne sont pas linéaires.
Par conséquent, les dangers liés à une variation de pH sont très différents selon le secteur dans l’échelle de ce type de mesure dans lequel le malade se trouve antérieurement à la variation de la concentration protonique.
3- Équilibre acide-base :
Selon le concept développé par Bronstedt, un acide est défini comme une substance capable de libérer un proton, et une base comme une substance capable d’accepter H+.
Un acide est ainsi toujours couplé à une base, selon l’équation :
acide <=> base + H+ (équation 2)
En physiologie et en physiopathologie, il est très important de différencier deux classes d’acides, l’acide carbonique (H2CO3) et les autres acides.
En effet, ces deux types d’acides diffèrent très notablement en ce qui concerne leur production journalière et leur voie d’élimination.
Chaque jour, l’organisme produit environ 15 000 mmol d’anhydride carbonique (CO2), qui vont se combiner à l’eau pour former de l’acide carbonique (H2CO3), lequel va se dissocier en libérant un proton, donc abaisser le pH, selon l’équation :
CO2 + H2O <=> H2CO3 <=> HCO3– + H+ (équation 3) où HCO3– : bicarbonate, la réaction étant catalysée, dans l’organisme, par l’enzyme anhydrase carbonique.
Le CO2 est éliminé par les poumons, ce qui prévient une accumulation de protons qui mènerait très rapidement au décès si cette voie d’excrétion n’était pas disponible.
Des acides non carboniques sont également produits quotidiennement, essentiellement à partir du métabolisme des protéines, mais en quantité beaucoup moins importante, soit environ 50 à 100 mmol/jour.
Ces acides vont être excrétés par les reins, au moins si l’on en croit la théorie dite classique.
4- Loi d’action de masse :
Le respect de la loi d’action de masse veut que la vitesse d’une réaction chimique soit proportionnelle au produit de la concentration des substances qui interagissent. Par exemple, dans le cas de l’eau qui se dissocie en hydrogène et hydroxyl, selon l’équation :
H2O Û OH– + H+ (équation 4) la vitesse (V1) de dissociation de l’eau est proportionnelle à sa concentration et peut s’écrire :
V1 = k1 [H2O] (équation 5) alors que la réaction chimique réciproque se déroule à une vitesse V2 qui est :
V2 = k2 [OH-] [H+] (équation 6) où k1 et k2 représentent les deux coefficients de proportionnalité des deux équations.
À l’équilibre des deux réactions de sens opposé, V1 = V2, donc :
K = k1/k2 = [OH–] [H+]/[H2O] (équation 7)
Cette relation établit donc, pour une substance chimique donnée (ici l’eau), quelles vont être les concentrations relatives des différentes composantes des réactions chimiques engagées.
La constante K décrit quantitativement cet équilibre.
La loi d’action de masse étant valide pour toute réaction chimique, rien n’empêche qu’elle soit appliquée aux couples acide-base, et ce selon l’équation :
Ka = [A–] [H+]/[HA] (équation 8) dans laquelle [A–] représente la base (un anion), et Ka la constante de dissociation, ou encore d’ionisation de l’acide considéré.
Les acides diffèrent par leur constante propre Ka.
Ainsi, certains acides sont, dans l’organisme et dans les limites compatibles avec la vie, très dissociés, ils ont donc un Ka élevé (acides forts), alors que d’autres sont peu dissociés et sont caractérisés par un Ka bas (acides faibles).
Le pKa n’est autre que le cologarithme du Ka.
Un acide qui présente un pKa inférieur au pH physiologique de 7,40, est majoritairement dissocié dans ces conditions (par exemple l’acide carbonique, pKa 6,10), alors que l’ammonium (NH4 +), qui est caractérisé par un pKa de 9,25, n’est pratiquement jamais présent sous forme dissociée dans les liquides biologiques compatibles avec la vie.
Un acide fort est donc caractérisé par un pKa bas, une base forte par un pKa élevé.
L’équation (8) peut être réarrangée comme suit :
[H+] = Ka [HA]/[A–] ou sous la forme logarithmique :
pH = pKa + log [A–]/[HA] (équation 9)
Cette nouvelle équation permet de décrire ce que pourrait être le pH d’une solution donnée contenant un acide avec un certain Ka, en fonction des concentrations respectives d’acide ([HA]) et de base ([A–]) dans le milieu considéré.
Rien n’empêche d’appliquer cette équation au couple acide-base/acide carbonique-bicarbonate.
L’équation devient alors :
pH = pKa + log [HCO3–]/[H2CO3] (équation 10) ou équation de Henderson-Hasselbach, appliquée à ce couple acidebase, pour lequel le pKa est de 6,10.
Comme il existe une relation entre l’anhydride carbonique (CO2) et l’acide carbonique (H2CO3), selon l’équation (3), on peut écrire :
pH = pKa + log [HCO3–]/0,03 PCO2 (équation 11)
En effet, le CO2 dissous en milieu aqueux est mesuré à l’aide de sa pression partielle (PCO2) et le coefficient de proportionnalité entre ces deux valeurs est précisément 0,03, pour autant que la PCO2 soit exprimée en mmHg.
5- Tampons :
Tous les couples acide-base sont capables d’accepter ou de rejeter des protons, selon les circonstances. Les acides faibles, peu dissociés dans les conditions physiologiques dans les milieux aqueux de l’organisme, agissent tantôt en captant, tantôt en libérant des H+.
Leur présence est donc à même d’atténuer les variations de concentration protonique qui pourraient survenir.
On les appelle, pour cette raison, des tampons.
Ceux-ci sont présents dans le milieu extracellulaire (essentiellement sous la forme du couple acide carbonique-bicarbonate) et dans le milieu intracellulaire (protéines, tissu osseux, etc).
La concentration protonique d’une solution aqueuse contenant plusieurs couples acide-base, dont des tampons, affecte chacun de ces couples, conformément à la loi d’action de masse :
[H+] = Ka1 [HA1]/[A1-] = Ka2 [HA2]/[A2–] = Kan [HAn]/[An–] (équation 12)
Ainsi, si le Ka et les concentrations relatives des composantes acide et basique d’un tampon donné et le pH de la solution sont connus, l’état de tous les tampons de la solution, pour autant qu’un équilibre ait été atteint, peut être calculé.
C’est le principe iosohydrique.
Son application clinique est importante : connaissant le pH d’un échantillon sanguin, ainsi que sa concentration en bicarbonate et la PCO2, l’état de l’ensemble des tampons de l’organisme peut être estimé de façon assez précise.
C’est l’équilibre des tampons, concept que nous retrouvons plus loin en discutant la notion d’excès ou de déficit de base.
Par ailleurs, l’étude du couple bicarbonateacide carbonique est particulièrement intéressante, car non seulement ses composants sont assez faciles à doser, mais encore chacun d’entre eux est l’objet d’une régulation particulière (ventilation pour l’acide carbonique-CO2 et élimination des acides fixes pour le bicarbonate).
B – PRODUCTION ET ÉLIMINATION DES PROTONS :
Nous avons vu que deux sources protoniques existent dans l’organisme, une source directe, liée au métabolisme, et une source indirecte, très abondante, liée à la production d’anhydride carbonique (CO2) qui va être éliminé par les poumons.
Ainsi, même si nos aliments sont le plus souvent neutres sur le plan acidobasique, des acides sont générés lors des processus cataboliques, soit normaux (acide pyruvique, lactique, acides du cycle tricarboxylique de Krebs), soit pathologiques (acides cétoniques, par exemple).
De plus, l’hydrolyse, réversible de l’adénosine triphosphate (ATP) est la principale source de H+ issus d’un organisme vivant, selon la réaction ci-dessous :
ATP4– + H2O <=> ADP3- + H+ + HPO4 2– (équation 13)
La production endogène d’acides fixes se monte donc, nous l’avons vu, à environ 50-100 mEq par jour, soit environ 1-1,5 mEq/kg de poids corporel par jour.
Selon la théorie classique, c’est le rein qui est chargé de l’élimination de ces acides, alors que, selon Atkinson, le foie joue un rôle essentiel dans cette élimination, via le cycle de l’urée et de l’ammonium.
Le fait que la production journalière d’acide carbonique est d’environ 15 000 mEq, alors que celle des acides fixes n’est que de 50-100 mEq/jour, explique pourquoi, en cas de défaillance respiratoire et d’hypoventilation, l’acidémie est rapide, alors qu’elle prend plusieurs jours pour s’installer en cas de défaillance rénale.
Il faut bien comprendre que, même si, lors des processus métaboliques, de grandes quantités de protons sont produites et consommées, la production nette de H+ est modeste, comme nous l’avons vu plus haut.
En effet, il existe un équilibre entre production de H+ et élimination de protons, chaque voie métabolique (glucides, lipides, protides) possédant une ou des voies métaboliques libérant des protons, et une ou des voies métaboliques consommant des protons.
Ainsi, les hydrates de carbone produisent des protons lors de la glycolyse, sous la forme d’acide lactique, un acide fort, de sorte que, chaque jour, environ 1 300 mmol de cet agent sont produites.
L’acide lactique est catabolisé par les tissus, surtout le foie, via le cycle de Cori.
Lors de ces opérations, la consommation d’un proton est obligatoire. Celle-ci empêche le développement d’une acidose, en l’absence d’accumulation pathologique de ces produits.
C’est ce qui explique que l’administration exogène de sels comme le lactate, ou l’acétate, mène à l’alcalémie, par consommation de H+ lors de leur catabolisme.
Les lipides, via la lipolyse, mènent à la production d’environ 300 mmol/jour de H+ chez le sujet normal, résultant de la dissociation des acides cétoniques.
La synthèse des triglycérides et des acides gras consomme la majorité des protons produits.
Quant aux protides, ils génèrent, chez le sujet normal avec un régime comprenant environ 100 g de protéines, à peu près 1 000 mmol/jour de H+ .
Ces protons proviennent essentiellement du foie, lors du processus de la synthèse de l’urée.
Cette production de H+ est toutefois largement contrebalancée par la consommation des protons qui est consécutive à l’oxydation complète des squelettes carbonés des acides aminés neutres.
C – THÉORIE DE PETER STEWART :
Nous avons vu plus haut que la concentration des liquides biologiques en protons ([H+]), en particulier celle du compartiment vasculaire, était très finement ajustée pour s’établir à 40 ± 5 nmol/L (pH 7,40 ± 0,05), et ce grâce aux équilibres métaboliques entre production et élimination des protons, d’une part, et au moyen des actions des poumons et des reins.
C’est la théorie dite classique du métabolisme acide-base, qui place au centre du débat la régulation de la concentration protonique dans les liquides biologiques.
Toutefois, avant d’envisager, puis de comprendre, comment l’organisme parvient à ce résultat étonnant par sa précision, il convient de scruter avec plus d’acuité quels sont les déterminants physicochimiques de la concentration [H+] dans un milieu aqueux.
1- Régulation de la concentration protonique :
Les liquides biologiques contiennent essentiellement de l’eau, une source virtuellement inépuisable de protons qui peuvent être générés par la dissociation de ce liquide.
Ce sont les lois de la chimie physique (principes de la thermodynamique) qui déterminent cette dissociation, en particulier le principe de l’électroneutralité (la somme des ions chargés négativement est égale à celle des ions positifs) et la loi de la conservation de la masse (la quantité de matière demeure constante, à moins que l’on ajoute ou génère, détruise ou retranche, une partie ou l’ensemble de la substance considérée).
Dans l’eau pure, le principe de l’électroneutralité veut que, en termes de concentration, [H+] = [OH–].
Dans des solutions plus complexes, comme le plasma, d’autres facteurs agissent sur la dissociation de l’eau, mais la source de protons demeure la dissociation de l’eau de façon extrêmement majoritaire.
En connaissant la valeur de trois facteurs agissant sur cette dissociation de l’eau, la PCO2, la différence des ions forts (ou strong ion difference [SID]) et la concentration totale des acides faibles, peu dissociés (Atot), on peut arriver à déterminer la valeur de la concentration [H+].
C’est ici que la théorie de Stewart diffère essentiellement de la théorie classique : [H+] ne varie, donc [HCO3 –] ne change, que si l’une ou plusieurs des trois variables que nous venons de décrire se modifie.
Ainsi, [H+] et [HCO3 –] ne sont plus des variables indépendantes, et leur taux dans les liquides biologiques va dépendre de la régulation, par l’organisme, des trois grandeurs mentionnées ci-dessus en obéissant aux lois physicochimiques de la thermodynamique.
C’est donc l’état physicochimique du milieu aqueux qui va déterminer maintenant la quantité de protons dans ce même milieu qui est susceptible, en fonction de cet état, de gérer ou d’absorber des ions [H+] en jouant avec la dissociation de l’eau.
Envisageons maintenant successivement les rôles de ces trois facteurs sur le pH, la concentration des ions [H+].
* pCO2 :
L’anhydride carbonique (CO2), en réagissant avec l’eau selon la réaction :
CO2 + H2O <=> H2CO3 <=> H+ + HCO3 – (équation 3) génère à la fois un proton et un anion bicarbonate.
L’augmentation du CO2 a donc bien accru de façon directe la dissociation de l’eau et, par ce mécanisme, provoqué une charge acide. Réciproquement, la diminution de la charge acide sera réalisée dans l’organisme par la ventilation alvéolaire.
* Différence des ions forts :
Les anions fortement dissociés jouent un rôle dans la dissociation de l’eau via le principe de l’électroneutralité.
Il existe une différence entre la somme des concentrations des cations fortement dissociés mesurés (essentiellement Na+, K+, Mg++ et Ca++) et celle des concentrations des anions fortement dissociés mesurés (Cl– et, selon les circonstances, les anions lactate, sulfate, acétoacétate, etc).
Cette différence est appelée strong anions difference apparente (SIDa). SIDa équivaut pratiquement à :
SIDa = ([Na+] + [K+] + [Mg++] + [Ca++]) – ([Cl–] + [lactate–]) (équation 14)
En l’absence de pathologie (lactate et sulfate normaux et en l’absence de corps cétoniques ou d’autres anions forts, etc), cette différence est de 40-42 mEq/L.
Il faut noter que ni H+ ni HCO3 – ne sont des ions forts, car leur dissociation est incomplète, ces ions s’associant facilement à d’autres ions, comme OH– pour le proton, par exemple, ou encore H+ pour HCO3 –.
Lorsque de nouveaux anions forts apparaissent, ou si le chlore augmente, par exemple, cette SIDa se réduit et le principe d’électroneutralité ne peut être respecté que par l’augmentation de la dissociation de l’eau, laquelle génère des protons pour augmenter la quantité de cations, et mène à l’acidose.
Inversement, une augmentation du SIDa réduit la dissociation de l’eau et mène à une alcalose.
SIDa basse => acidose
SIDa élever => alcalose
Toutefois, en raison de la non-linéarité de la relation entre la modification du SIDa et l’importance de la dissociation de l’eau, une augmentation du SIDa au-dessus de sa valeur normale de 40 mmol/L a un fort effet alcalinisant, alors qu’un abaissement équivalent (équimolaire) du SIDa a un effet acidifiant moindre.
* Concentration totale des acides faibles :
Les acides faibles peu dissociés, représentés surtout par le phosphate et par les protéines, sont présents, par définition puisqu’ils sont peu dissociés, sous les formes A– (dissociée) et AH (protonée), l’ensemble étant appelé Atot, où [Atot] = [A–] + [AH].
En fait, l’équilibre entre [A–] et [AH] dépend de la PCO2 et des altérations de SIDa, et ne peut donc pas être considéré comme une variable indépendante intervenant sur la dissociation de l’eau.
C’est bien plutôt [Atot] qui représente la troisième variable indépendante agissant sur cette dissociation. Les concepts développés par Stewart sont une véritable révolution copernicienne dans notre approche du métabolisme acidobasique.
Toutefois, ils sont loins d’avoir acquis un droit de cité en clinique, où l’approche classique reste bien implantée, malgré son caractère conceptuellement insatisfaisant.
La complexité de l’approche de Stewart va continuer à faire les beaux jours de la théorie classique.
Perturbations de l’équilibre acide-base :
Une perturbation de l’équilibre acide-base peut être due à une altération de la fonction ventilatoire (acidose ou alcalose respiratoire, en cas d’hypo- ou d’hyperventilation alvéolaire), ou encore lorsqu’une charge acide ou alcaline dépasse les possibilités d’élimination de l’organisme (acidose ou alcalose métaboliques).
Les termes d’acidémie et d’alcalémie sont employés quand le pH du liquide examiné, le plus souvent le sang artériel, est acide ou alcalin.
Les termes d’acidose et d’alcalose sont réservés à la description du ou des processus qui tendent à abaisser ou à élever le pH.
Le plus souvent une acidose s’accompagne d’acidémie et, réciproquement, una alcalose d’alcalémie.
Toutefois, lorsqu’un malade présente simultanément plusieurs perturbations acidobasiques (troubles acidobasiques complexes), le pH qui résulte de ces troubles va dépendre de leur importance relative.
Ces troubles acidobasiques sont alors dits mixtes ou combinés.
A – ACIDOSE RESPIRATOIRE :
1- Physiopathologie de l’acidose respiratoire :
Si la PaO2 artérielle diminue avec l’âge, en raison de l’altération des propriétés mécaniques des poumons (perte des fibres élastiques pulmonaires, à l’origine de la génération de zones à rapport ventilation-perfusion abaissé), la PaCO2, dès l’âge de 3 ans est constante (40 ± 5 mmHg ; 5,3 ± 1,0 kPa) chez le sujet normal vivant à basse altitude.
La PaCO2, c’est-à-dire le taux de CO2 dissous dans le sang, le plasma ou les tissus, est le résultat d’un équilibre entre la production de CO2 (VCO2) et son élimination, la ventilation alvéolaire (VA), tel que l’équation ci-dessous le décrit :
PaCO2 = k V? CO2/VA (équation 15)
où k est une constante permettant d’ajuster les unités de mesure (kPa pour la PaCO2, mL/min pour la VCO2 et L/min pour VA), alors que l’équation ci-dessous décrit les facteurs qui déterminent VA :
VA = fr (VT – VD) (équation 16)
où fr : fréquence respiratoire, VT : volume courant, VD : volume de l’espace mort.
Ainsi, une hyper- ou une hypocapnie sont-elles le résultat d’un déséquilibre entre la production de CO2 et son élimination.
Une hypercapnie est donc consécutive soit à une augmentation de la VCO2 sans adaptation de la ventilation alvéolaire, ou à une baisse de la ventilation alvéolaire, elle-même pouvant résulter d’une baisse de la fréquence respiratoire ou du volume courant, ou à une élévation de l’espace mort (VD), ou à une combinaison de ces altérations.
Une conséquence de l’hypercapnie est l’apparition d’une hypoxémie, comme le prédit l’équation de l’air alvéolaire :
PaO2 = PAO2 – DA-aO2 (équation 17)
où : PAO2 : pression partielle d’O2 alvéolaire et DA-aO2 : différence alvéoloartérielle en O2, la PAO2 étant calculée selon l’équation :
PAO2 = (PB – PH2O) FIO2 – PaCO2/R (équation 18)
où PB : pression atmosphérique (760 mmHg au niveau de la mer), PH2O pression de vapeur d’eau à saturation complète, soit 47 mmHg (6,3 kPa) ; FIO2 : fraction inspirée d’O2 ; PaCO2 : pression partielle d’anhydride carbonique artérielle ; R : quotient respiratoire, généralement fixé à 0,7.
La deuxième conséquence de l’hypercapnie est la survenue d’une acidémie.
En effet, si l’on considère l’équation (3), il est évident que toute augmentation du CO2 dissous va générer l’apparition d’un H+, donc abaisser le pH, et vice-versa.
Cette équation explique aussi le fait qu’il est impossible, pour des raisons purement physicochimiques, de faire varier la quantité de CO2 dissous dans un milieu aqueux, comme le sont les liquides intra- et extracellulaires de l’organisme humain, sans faire varier, dans le même sens, le taux de bicarbonate.
Ainsi, le taux « normal » de bicarbonate plasmatique de 24 ± 1 mmol/L n’est-il situé à cette valeur dans le liquide extracellulaire que si la PCO2 est elle-même normale, c’est-à-dire située autour de 5,3 ± 0,5 kPa (40 mmHg).
L’adéquation entre le taux de bicarbonate mesuré dans échantillon de sang et la PCO2 est rendue par le concept d’ « excès de base ».
Si le taux de bicarbonate mesuré est trop élevé par rapport au niveau de PCO2, selon ce que la dissociation de l’eau peut prédire (équation 3), il y a un « excès de base » (base excess > 0,0 mmol/L), si ce taux est inférieur à la valeur attendue, il y a « déficit de base » (base excess < 0,0 mmol/L).
* Acidose respiratoire aiguë :
En cas d’acidose respiratoire aiguë, c’est-à-dire dans la situation où l’élévation de la PaCO2 survient avant que les mécanismes de compensation rénale n’aient eu le temps de se mettre en route (moins de 12-24 heures), il n’y a pas d’excès de base, mais un excès de protons dans le sang et le reste du liquide extracellulaire, ce qui est rendu par une baisse du pH, et une ascension du taux de bicarbonate.
Cette ascension du taux de bicarbonate excède de loin en quantité celle du taux de proton, bien que l’équation (3) prédise une augmentation équimolaire de ces deux ions quand la PCO2 monte.
En effet, comme mentionné plus haut, le taux du proton dans le plasma est très bas (40 nEq/L à l’état normal), et une ascension même faible de la concentration de H+ est rapidement létale.
La simple augmentation de PaCO2 de 1,0 kPa, qui passerait ainsi de 5,3 à 6,3 kPa (40 à 48 mmHg), une situation pourtant fréquente, serait incompatible avec la vie, le pH devenant très inférieur à la valeur de 6,8 considérée comme la plus basse valeur non létale.
Le proton généré dans le milieu extracellulaire diffuse rapidement vers le milieu intracellulaire, ce qui n’est pas le cas pour le bicarbonate.
Ceci prévient une baisse massive du pH extracellulaire.
Dans les cellules, l’ion H+ est tamponné par des bases (sulfates, phosphates, protéines, etc).
Dans cette situation, il existe une augmentation de la quantité des bases extracellulaires (le bicarbonate) en même temps que survient une consommation, dans la même mesure, des bases intracellulaires.
En considérant l’ensemble de l’organisme, ici un individu donné formé de cellules et de liquide extracellulaire, il ne se produit qu’un transfert de bases, le bilan net étant à zéro pour l’individu entier.
Les taux des bases extracellulaires et les bases intracellulaires sont diminués de la même façon.
Il n’y a donc aucun « excès de base », c’est-à-dire qu’aucune base n’a été ajoutée à l’ensemble de l’organisme (liquides intra- et extracellulaires), mais un simple déplacement d’alcalis (gain de bases extracellulaires et perte de bases intracellulaires).
En moyenne, en acidose respiratoire aiguë, on estime que le taux plasmatique de bicarbonate s’élève d’environ 0,75 mmol/L pour 1 kPa (7,5 mmHg) d’élévation de la PaCO2.
* Acidose respiratoire chronique :
Lorsque l’hypercapnie persiste (acidose respiratoire chronique), notamment plusieurs jours, il apparaît une compensation rénale.
Les cellules tubulaires rénales proximales activent l’antiport Na+-H+ et l’on assiste à une sécrétion augmentée d’acide au niveau distal, associée à une résorption accrue de bicarbonate, ce qui ramène le pH plasmatique à un taux proche de sa valeur normale, en 3 à 5 jours si l’hypercapnie persiste.
Simultanément à l’augmentation du taux sanguin de bicarbonate, la chlorémie diminue, en partie parce que cet anion est échangé contre le bicarbonate au niveau des globules rouges, et en partie parce que la chlorurie augmente.
Il en résulte un état d’hyperbicarbonatémie hypochlorémique, qui peut être à l’origine, une fois l’hypoventilation corrigée et la PaCO2 normalisée, de la persistance d’un taux de bicarbonate élevé (alcalose posthypercapnique), qui ne pourra être normalisé que par l’adjonction de sels de chlore (KCl, NaCl) ou d’acétazolamide.
En moyenne, en acidose respiratoire chronique, le taux plasmatique de bicarbonate s’élève de 4,0 mmol/L pour 1 kPa (7,5 mmHg) d’élévation de la PaCO2.
2- Causes de l’acidose respiratoire :
La classification des causes d’acidose respiratoire peut découler des paramètres décrits par l’équation vue plus haut :
PaCO2 = k VCO2/VA (équation 19)
* Acidoses respiratoires causées par une augmentation de la production de CO2 (VCO2) :
La production de CO2 est le résultat de l’activité métabolique.
En état régulier, au repos, l’organisme produit environ 3-4 mL de CO2/kg de poids corporel (VCO2).
L’effort physique, la fièvre (il existe une augmentation de la VCO2 de 13 % par degré centigrade d’accroissement de température corporelle), une hyperthyroïdie ou un état catabolique, comme dans le sepsis, accroissent la VCO2.
Le type de nutriment ingéré modifie lui aussi la production de CO2. Par exemple, le catabolisme lipidique mène à un quotient respiratoire (QR) de 0,7, alors que le catabolisme purement glucidique conduit à un QR de 1,0.
Pour mémoire, le quotient respiratoire, QR, est défini par l’équation ci-dessous :
QR = VCO2/VO2 (équation 20)
où VCO2 : production de CO2, VO2 : consommation d’O2 (normale : 2-4 mL/kg de poids corporel au repos).
Il est rare qu’une augmentation de la VCO2 mène à une hypercapnie, donc à une acidose respiratoire.
En effet, la plupart du temps, l’organisme peut augmenter en parallèle VCO2 et VA, ce qui maintient une PaCO2 normale.
En revanche, en cas de limitation ventilatoire, comme dans les neuromyopathies affectant les muscles respiratoires, ou la bronchopneumopathie chronique obstructive (BPCO), la ventilation peut être limitée et une augmentation de la VCO2 peut mener à une hypercapnie.
Il en va de même si un patient est soumis à la ventilation mécanique en présence d’une sédation profonde ou de myorelaxants : il ne peut alors faire face à une augmentation de la VCO2, puisque sa ventilation est entièrement dépendante du respirateur.
Enfin, en cas de limite ventilatoire, l’administration de quantités exagérées d’hydrates de carbone pourrait causer une hypercapnie.
Ce facteur a parfois été incriminé dans le sevrage difficile du respirateur mécanique.
* Acidoses respiratoires consécutives à une diminution de la ventilation alvéolaire (VA) :
+ Diminution de la ventilation-minute (VE = FB × VT) :
Le malade est dans ce cas incapable, totalement, ou partiellement, de mettre en mouvement ses muscles inspiratoires (« le malade ne veut pas respirer ») car la commande respiratoire centrale, neurologique, est déficiente (atteinte des centres respiratoires bulbopontiques).
L’origine de ce déficit peut être variable.
L’autre possibilité veut que la genèse de la commande respiratoire neurologique est normale, mais qu’elle n’est pas transmise aux muscles inspiratoires (atteinte des faisceaux neurologiques, des cornes antérieures de la moelle, des nerfs périphériques ou de la jonction neuromusculaire), ou encore que le malade envoie bien un signal à ses muscles inspiratoires, mais que ceux-ci n’y répondent pas (myopathies, fatigue musculaire respiratoire, ou encore impossibilité totale ou partielle de mouvoir un système respiratoire passif trop anormal, par exemple en cas de cyphoscoliose causant une rigidité de la cage thoracique anormale).
+ Augmentation de l’espace-mort :
Une hypoventilation peut être causée par une augmentation du VD.
En général, lorsque la pompe ventilatoire est normalement active, l’effet d’une augmentation du VD est largement compensé par une augmentation de la ventilation-minute, car les centres respiratoires n’autorisent pas une élévation de la PaCO2 qui serait la conséquence d’une hypoventilation.
En revanche, lorsque cette même pompe est défaillante, l’effet de VD se démasque et la PaCO2 s’élève.
Un exemple typique de cette situation, fréquent en clinique, est l’apparition d’une respiration rapide et superficielle lorsque les muscles inspiratoires se fatiguent.
Une hypercapnie survient alors.
3- Conséquences de l’acidose respiratoire :
La conséquence la plus dangereuse de l’acidose respiratoire est liée à l’apparition de l’hypoxémie et de ses effets, notamment sur le cerveau.
L’hypercapnie, quant à elle, a essentiellement des effets neurologiques (tremor, astérixis, somnolence, agitation, angoisse, délire et, pour des PaCO2 très élevées, coma, notamment).
Le volume sanguin cérébral s’accroît en raison d’une vasodilatation cérébrale, pouvant mener à l’oedème cérébral. Le système cardiovasculaire est lui aussi affecté par l’hypercapnie.
Le taux des catécholamines circulantes est augmenté, ce qui provoque une tachycardie, mais l’effet direct du CO2 sur les muscles lisses artériolaires mène à une vasodilatation, le tout aboutissant à un état hyperdynamique.
La peau est chaude, rouge, le malade transpire.
L’hypercapnie aiguë sévère peut conduire à une baisse de la contractilité cardiaque, probablement induite par l’acidose intracellulaire, laquelle interfère avec le couplage du calcium et de l’actine-myosine.
De plus, l’hypercapnie diminue l’affinité de l’hémoglobine pour l’oxygène (effet Bohr), ce qui favorise l’extraction de l’oxygène en périphérie.
L’acidose respiratoire potentialise la vasoconstriction hypoxique pulmonaire et, mais ce point est controversé, elle aurait un effet négatif sur la contractilité musculaire squelettique, notamment au niveau des muscles respiratoires.
L’hypercapnie chronique peut aboutir à la rétention hydrosaline et à l’apparition d’oedèmes, sans que le débit cardiaque ne soit abaissé.
La vasodilatation liée à la rétention de CO2 joue un rôle central dans cette rétention sodique.
* Hypoventilation contrôlée (permissive hypercapnia) :
Cette stratégie thérapeutique de réanimation consiste, en ventilation mécanique, à protéger le système respiratoire des barotraumatismes en réduisant la ventilation, laissant alors la PaCO2 augmenter de façon réglée, quitte à corriger le pH au moyen d’un tampon.
Cette technique de réanimation est employée, soit dans les syndromes obstructifs sévères (asthme, BPCO), soit dans le syndrome de détresse respiratoire aigu.
Pour autant que le malade ne présente pas initialement d’atteinte cérébrale à même de générer une hypertension intracrânienne, et pour autant que l’ascension de la PaCO2 ne soit pas trop rapide et trop élevée, cette technique paraît sûre.
Certains auteurs ont récemment suggéré que l’hypercapnie per se pourrait exercer des effets bénéfiques au niveau pulmonaire myocardique et cérébral, mais l’importance clinique de leurs travaux reste à démontrer.
4- Traitement de l’acidose respiratoire :
Il est essentiellement dirigé vers la cause de l’acidose (traitement d’une crise d’asthme, d’une intoxication médicamenteuse, par exemple).
En cas d’acidémie sévère et/ou d’échec des mesures thérapeutiques spécifiques à la cause de l’acidose, on doit recourir à la ventilation mécanique, invasive ou non.
En cas d’acidose respiratoire chronique, le bicarbonate plasmatique est élevé en raison de la compensation rénale.
Il faut veiller à ne pas provoquer une alcalose de reventilation au moyen d’une ventilation mécanique trop vigoureuse ; le cas échéant, l’élimination du bicarbonate en excès peut être accélérée au moyen d’acétazolamide.
L’administration d’O2 permet le plus souvent de corriger aisément l’hypoxémie due à l’hypoventilation.
Toutefois, chez les malades atteints de BPCO, il faut prendre garde à l’élévation de la PaCO2 sous O2, quel qu’en soit le mécanisme, et on met en route une oxygénothérapie contrôlée si le malade n’est pas placé en ventilation mécanique.
Quelques autres points sont également à noter. Tout d’abord, il convient d’éviter tout facteur à même d’entraver la fonction des muscles respiratoires (hypokaliémie, hypophosphatémie, hypomagnésémie et hypocalcémie, anémie).
Ensuite, on réalise que l’administration d’agents alcalinisants est potentiellement dangereuse, puisque, en l’absence de ventilation mécanique à même de régler la PaCO2, le bicarbonate une fois tamponné, va amener une charge supplémentaire en CO2, une charge osmotique considérable et il va acidifier le milieu intracellulaire ; l’administration de tris-hydroxyméthyl-aminométhane (THAM), en théorie, est un agent alcalinisant capable de réduire la PaCO2, mais cet effet n’est que modeste.
En effet, le THAM interagit avec le CO2 et le proton H+ selon les réactions suivantes :
THAM-NH2 + H+ <=> THAM-NH3 + THAM-NH2 + H2O + CO2 <=> THAM-NH3 + + HCO3 – (équations 21)
Enfin, les apports nutritionnels ne doivent pas être excessifs, notamment les apports glucidiques, comme nous l’avons vu plus haut.
B – ALCALOSE RESPIRATOIRE :
1- Physiopathologie de l’alcalose respiratoire :
Cette perturbation acidobasique est caractérisée par une diminution de la PaCO2, consécutive à une hyperventilation et assortie à un abaissement variable du taux plasmatique de bicarbonate, donc à une élévation du pH.
Il existe ainsi, dans cette situation, un déséquilibre entre la production de CO2 (VCO2) et son élimination par les poumons, et ce en faveur de cette dernière.
* Alcalose respiratoire aiguë :
Dans les 10 minutes qui suivent l’apparition de l’hyperventilation, les protons sortent du milieu intracellulaire et la réaction suivante se produit :
H+ + HCO3 => H2CO3 => CO2 + H2O (équation 3)
ce qui aboutit à un abaissement modéré du taux de bicarbonate plasmatique, une caractéristique de l’alcalose respiratoire aiguë.
Cet abaissement du taux de bicarbonate peut être calculé comme suit : en multipliant par 0,1 la variation en mmHg de la PaCO2, la valeur de l’abaissement du taux de bicarbonate plasmatique en mmol/L est obtenue. Par ailleurs, le taux plasmatique de lactate augmente, pour plusieurs raisons.
D’une part, l’enzyme phosphofructokinase (PFK) qui lance la glycolyse est activée, et la formation d’acide pyruvique, donc d’acide lactique, est augmentée.
D’autre part, l’excrétion de lactate des cellules dans lesquelles cet anion est produit vers le plasma est accrue, alors que le captage par le foie, le site de catabolisme du lactate, se voit réduit.
Toute hyperventilation aiguë se traduit donc par une hyperlactatémie modérée (variation de 1-3 mmol/L), sans acidémie.
En effet, le proton généré dans les cellules par la dissociation de l’acide lactique, un acide fort, est pris en charge par la chaîne respiratoire où il est oxydé et transformé en eau.
Enfin, plusieurs électrolytes sont affectés par l’alcalémie. D’une part, le calcium ionisé s’abaisse en raison du pH alcalin, ce qui peut mener à la tétanie d’hyperventilation.
Le potassium plasmatique s’élève transitoirement, sous l’effet de la stimulation adrénergique qui accompagne l’alcalose respiratoire, mais assez rapidement, la kaliémie chute, en raison d’un échange transmembranaire.
Le proton quitte la cellule, alors que le potassium se déplace vers le liquide intracellulaire. On estime que pour chaque variation de pH de 0,1 unité, on attend une diminution de 0,3 mEq/L de K+.
Quant à l’anion phosphate, lors de l’hyperventilation aiguë, en raison de l’élévation du pH, il pénètre lui aussi dans les cellules et on assiste à l’apparition d’une hypophosphatémie.
* Alcalose respiratoire chronique :
Elle se développe lorsque l’hyperventilation persiste et est caractérisée par une baisse plus marquée du taux de bicarbonate plasmatique.
Une estimation permet de dire que la diminution de bicarbonate plasmatique peut être calculée en multipliant par 0,4 la baisse de la PCO2, mesurée en mmHg.
Ici, le bicarbonate diminue en raison de la compensation rénale du trouble primaire respiratoire, le rein réagissant en réduisant la sécrétion tubulaire de H+, ce qui équivaut à diminuer la réabsorption du bicarbonate filtré.
Cet effet commence déjà 2 heures après le début de l’hyperventilation, mais il met 2 à 3 jours pour atteindre son maximum.
L’hyperventilation chronique mène, contrairement à l’hyperventilation aiguë, à l’hyperphosphatémie, en raison de l’apparition d’une résistance à l’action de l’hormone parathyroïdienne, ce qui conduit à une hypocalcémie et à une hypophosphaturie.
Cette effet semble dû à l’hypocapnie plutôt qu’à l’alcalémie, et il est en relation avec la stimulation de récepteurs b-adrénergiques.
2- Causes de l’alcalose respiratoire :
À part la situation artificielle de la ventilation mécanique, où c’est le thérapeute et non les centres respiratoires qui gouvernent la respiration, une hyperventilation peut être consécutive à la stimulation périphérique (présence de maladies respiratoires, via les fibres C et/ou les fibres J, ou via les chémorécepteurs en cas d’hypoxémie, notamment) ou à une stimulation centrale des centres qui règlent la ventilation (toxiques, fièvre, sepsis, etc).
3- Conséquences de l’alcalose respiratoire :
L’alcalose respiratoire provoque une augmentation de l’excitabilité des systèmes nerveux central et périphérique.
Ceci conduit à des symptômes comme des vertiges, des paresthésies, des crampes, des altérations de la conscience, parfois des convulsions.
De plus, l’alcalémie, via l’hypokaliémie en partie, mène à des arythmies cardiaques (tachyarythmies ventriculaires et supraventriculaires).
Chez des sujets prédisposés, l’alcalémie peut entraîner des douleurs thoraciques, soit par insuffisance coronarienne (vasospasmes coronariens) ou par spasme oesophagien.
Ces phénomènes se rencontrent surtout lorsque l’alcalémie est sévère (PaCO2 < 30 mmHg), et ils sont dus en partie à des altérations des potentiels de membrane et, en partie, à une vasoconstriction artérielle cérébrale.
4- Traitement de l’alcalose respiratoire :
En règle générale, l’alcalémie ne se traite pas en tant que telle, mais on traite sa cause.







")



{kind=link}